
O boală nelipsită din culturile de orz, pătarea reticulară

Pătarea reticulară a frunzelor de orz, o boală periculoasă, este nelipsită din culturile de orz, fiind prezentă an de an, cu frecvențe și intensități de atac mai ridicate sau mai scăzute, în funcție de condițiile climatice.
Vremea umedă din această perioadă susține infecțiile și dezvoltarea bolii. Scopul acestui articol tehnic este de a veni în sprijinul fermierilor cu date despre simptomatologia, biologia și strategia de combatere a acestei boli, care este periculoasă deoarece poate produce pagube importante în producția de orz și orzoaică.
Importanța economică
Pătarea reticulară este o boală foliară importantă a orzului, care poate produce pagube în producție, mai ales în zonele unde cad mai multe precipitații. Pe lângă reducerea producțiilor, calitatea boabelor are de suferit. În anii favorabili infecțiilor, pagubele în producție pot trece de 40% [Weibull et al., 2003; Backes et al., 2021]. Pagubele pot fi diferite, în funcție de rezistența soiurilor cultivate, virulența patogenului și condițiile climatice [Liu et al., 2011].

Recunoașterea simptomelor
Patogenul atacă frunzele, tulpinile, glumele, paleele, aristele și semințele. Pe frunze apar două tipuri de simptome, produse de forme diferite ale fungului:
-
Pete cu aspect de rețea sau reticulare (uneori arată ca ochiurile unui gard de sârmă) produse de Pyrenophora teres f. sp. teres. Aspectul reticular este dat de forma alungită a petelor, care arată ca niște striații și de dispunerea longitudinală și transversală a acestora. La început aceste striații au culoare gălbuie iar mai târziu brună - închis [Lightfoot & Able, 2010]. Astfel de pete pot fi observate toamna și primăvara devreme pe tinerele plăntuțe (observație personală);
-
Pete de culoare brună închis, de forme diferite, circulare și chiar eliptice produse de Pyrenophora teres f. sp. maculata. De jur împrejur, petele prezintă halou clorotic sau galben [Liu et al., 2011; Murray et al., 2009]. La atacuri puternice frunzele se necrozează și mor. După Tekauz (1985), zona galbenă sau clorotică din jurul petelor contează foarte mult în tabloul simptomatic al bolii, deoarece poate cuprinde întreaga frunză care în consecință se va ofili. Intensitatea atacului este corelată mai mult cu dimensiunea țesuturilor îngălbenite decât cu cele necrozate.
Deși cele două forme de Pyrenophora teres sunt foarte asemănătoare din punct de vedere morfologic, simptomele produse sunt diferite. După Akhavan et al. (2016), cele două forme sunt filogenetic distincte (considerate populații autonome genetic).

Pe lângă frunze, patogenul poate infecta și spicele. Când infecția ajunge la spic, pe glume și palee se formează pete liniare de culoare maroniu - închis. Atunci când bracteele florale sunt infectate este semn că patogenul a ajuns la semințe. Petele de pe semințe sunt de culoare mai închisă, aproape negre [Iacob et al., 1998; Popescu, 2005; Hatman et al., 1989]. După Comes et al. (1982), embrionul cariopselor infectate are culoare galben - brună, puterea germinativă fiind scăzută în astfel de situații.
De reținut că, la soiurile rezistente petele nu evoluează, rămânând mici. De regulă, nu prezintă halou galben sau clorotic decât excepțional. La soiurile sensibile, petele se extind crescând în dimensiuni. De jur împrejur, aceste pete au o zonă galbenă [Smedegard-Petersen, 1971; Tekauz, 1985].
De multe ori simptomele produse de Pyrenophora teres pot fi confundate cu cele produse de fungul Cochliobolus sativus. De aceea, diagnosticul trebuie confirmat pria analize de laborator.

Realizarea infecțiilor
Infecțiile pot apărea în culturile de orz încă din toamnă. Primele infecții pot fi din cauza miceliului din semințe, ascosporilor eliberați de ascele din peritecii, cât și conidiilor care iernează pe resturile culturilor infectate. În timpul vegetației, conidiile produc infecții secundare repetate [Popescu, 2005].
Supraviețuirea patogenului peste iarnă
Pyrenophora teres supraviețuiește pe resturile vegetale ale vechii culturi sub formă de micelii și pseudotecii brune [Smedegard - Petersen, 1972; Popescu, 2005]. Foarte importantă este supraviețuirea patogenului în semințele bolnave (inocul primar), fie la suprafața lor, fie sub pericarp [Singh & Chand, 1985].
Înmulțirea patogenului pe samulastra de orz și pe poacee sălbatice duce la un plus de inocul [Brown et al., 1993).
Condiții climatice favorabile infecțiilor
Patogenul se dezvoltă foarte bine în perioadele caracterizate de umezeală (sursa de inocul crește) - [Mclean et al., 2009].
Infecțiile cele mai periculoase la orz au loc atunci când umezeala persistă pe frunze 10 până la 30 de ore și chiar mai mult. În astfel de condiții, când umiditatea este aproape de maxim (100%) și temperaturile sunt cuprinse între 15 și 25°C (optim 22°C), sporularea ciupercii este maximă. În timpul sporulării petele capătă aspect catifelat, semn că s-au format conidioforii și conidiile ciupercii [Chuhina, 2008]. Precipitațiile continue favorizează infecțiile masive [Jordan, 1981].
Managementul integrat
În cadrul sistemului de combatere integrată există o serie de măsuri care trebuie utilizate echilibrat pentru a ține sub control această helmintosporioză. În ultimii ani, Pyrenophora teres a câștigat teren, fiind tot mai prezentă în culturile de orz (au crescut suprafețele cultivate). Semințele de orz, deși certificate, se pare că sunt infectate totuși, altfel nu se explică incidența și severitatea bolii tot mai ridicate, încă din toamnă. De aceea, se recomandă îmbinarea măsurilor profilactice cu cele chimice și biologice pentru ținerea sub control a pătării reticulare.
Factorii care susțin infecțiile sunt:
-
Rotațiile scurte;
-
Lucrările minimale ale solului;
-
Semănatul devreme;
-
Densitatea mare a plantelor;
-
Rezistența la fungicide;
-
Soiurile sensibile;
-
Precipitațiile continue [Locke, 1982; Gupta și Loughman, 2001; Doyle, 2005; Popescu, 2005].
Măsuri profilactice
Pentru succes în combatere și prevenirea daunelor, respectarea măsurilor profilactice este esențială.
Se recomandă:
-
Utilizarea soiurilor rezistente;
-
Utilizarea semințelor sănătoase, certificate este esențială în prevenție, deoarece pe și în semințe, patogenul poate fi prezent. După Youcef-Benkada et al. (1994), semințele bolnave pot contribui la introducerea patogenului în zone unde acesta nu era prezent;
-
Evitarea monoculturii;
-
Rotația culturilor. Pentru a preveni infecțiile, rotația ar trebui să fie de minim doi ani între culturile de orz [Duczek et al., 1999];
-
Distanța dintre culturile noi și vechi ar trebui să treacă de 500 m, pentru a preveni infecțiile realizate de ascosporii și conidiile aduse de vânt;
-
Distrugerea samulastrei;
-
Tehnologii convenționale de cultură care se bazează pe lucrările solului;
-
Respectarea epocii optime de semănat. În cazul acestui patogen, semănatul timpuriu favorizează infecțiile. Într-un studiu publicat în 1987, Delserone & Cole arată că, în culturile semănate devreme, intensitatea atacului a fost mult mai ridicată iar în cele semănate mai târziu a fost mai scăzută;
-
Eliminarea sau reducerea sursei de inocul din câmpuri [Popescu, 2005; Liu et al., 2011]. Sursa de inocul poate fi redusă prin arături la 15 - 20 cm. Această lucrare este necesară deoarece pseudoteciile își pot păstra viabilitatea timp de aproape 9 luni iar microscleroții 2 ani [Piening, 1967; Ali - Haimoud et al., 1993];
-
Fertilizarea echilibrată cu azot. Este cunoscut că azotul favorizează dezvoltarea patogenilor în general, deoarece duce la creșterea umidității în interiorul culturii [Iordan & Hutcheon, 1984; Popescu, 2005].

Măsuri chimice
În cadrul strategiei de combatere, măsurile chimice dețin ponderea în prezent. Tratarea semințelor și aplicările foliare în timpul sezonului de vegetație sunt importante în controlul pătării reticulare a orzului.
Tratarea semințelor este esențială. Fungicidele omologate în țara noastră pentru tratarea semințelor sunt: Fludioxonil + protioconazol + tebuconazol; Protioconazol + tebuconazol; Difenoconazol + fludioxonil + tebuconazol; Fludioxonil + triticonazol; Tebuconazol; Fluxapyroxad; Triticonazol [după aplicația Pesticide 2.25.12.1, 2026].
Tratamentele în vegetație trebuie făcute la momentele optime, ținându-se cont și de pragul economic de dăunare (PED). În cazul pătării reticulate, PED-ul este de 20% intensitate de atac la începutul înspicării. Scopul tratamentelor este de a menține libere de patogen primele trei frunze, mai ales în timpul umplerii boabelor. La orz, în mod special, frunzele 2 și 3 sunt foarte importante pentru viitoarea producție (la grâu este importantă frunza stindard) - Weppler & Hollaway, 2004; Popescu, 2005].
Momente optime de aplicare a fungicidelor sunt: la apariția ultimei frunze (stindardul), precum și la începutul înspicării când PED-ul este de 20% intensitate.
În primăverile umede, când patogenul se instalează devreme în culturi se pot face tratamente și la începutul alungirii paiului (GS 30 - 33). Acest tratament protejează de infecție frunza 2 (prima de sub steag) care se implică în producție într-un procent cuprins între 20 - 40%. Tratamentul de la apariția primelor spice (GS 49) protejează frunza 3 care se implică în producția finală în procent de 10 până la 15% [Dunne, 2002]. Frunza steag sau stindardul este implicată cel mai puțin în producția finală (5 - 9%). Tratamentele făcute în stadii timpurii (GS 12) nu și-au dovedit eficacitatea.
În anii cu infecții masive poate fi necesar și un tratament în stadiul GS 58 pentru a reduce severitatea bolii.
Fungicidele omologate în România pentru combaterea pătării reticulare a orzului (dar și pentru alți patogeni ai cerealelor) sunt: Azoxistrobin; Azoxistrobin + protioconazol; Bixafen +spiroxamină + trifloxistrobin; Protioconazol + tebuconazol; Benzovindiflupir + protioconazol; Benzovindiflupir; Metrafenonă; Azoxistrobin + difenoconazol + tebuconazol; Protioconazol + trifloxistrobin; Protioconazol + spiroxamină + trifloxistrobin; Bixafen + tebuconazol; Difenoconazol; Fenpropidin; Piraclostrobin; Fluxapiroxad + piraclostrobin; Fluxapyroxad + metconazol; Fluxapyroxad + mefentrifluconazol; Mefentrifluconazol + piraclostrobin; Mefentrifluconazol; Mefentrifluconazol + metrafenonă + piraclostrobin; Mefentrifluconazol + protioconazol; Metconazol; Protioconazol; Tebuconazol; Tebuconazol + trifloxistrobin; Bixafen + protioconazol; Boscalid + kresoxim metil; Proquinazid + protioconazol; Folpet; Folpet + protioconazol; Azoxistrobin + folpet; Fluxapyroxad + protioconazol; Bixafen + protioconazol + spiroxamină.
nAzoxistrobin + difenoconazol + tebuconazol; Azoxistrobin + fluxapyroxad; Difenoconazol + protioconazol; Difenoconazol + metconazol; Fenpicoxamida + protioconazol; Kresoxim - metil + mefentrifluconazol [după aplicația PESTICIDE 2.25.12.1, 2026].
Măsuri biologice
Agenții biologici pot fi introduși în strategiile de management actuale ale pătării reticulare a orzului, mai ales la tratarea semințelor (în fermele ecologice cât și în cele convenționale). Atunci când se utilizează agenții biologici, fermierii trebuie să respecte cu strictețe măsurile de prevenție pentru a avea succes în combatere. Introducerea unui tratament biologic în sistemele de combatere integrată poate diminua poluarea mediului și reziduurile de fungicide ce pot rămâne în cariopse, mai ales atunci când nu se respectă timpii de pauză.
Interes există în prezent pentru mai mulți agenți de biocontrol: Trichoderma sp., Clonostachys rosea izolat IK726, Pseudomonas chlororaphis MA 342, Bacillus subtilis etc [Copping, 2004; Jensen et al., 2016; Moya et al., 2018].
În anul 2004, Pseudomonas chlororaphis MA 342 era comercializat sub numele de Cedomon (BioAgri AB, Stockholm, Suedia) - Anon., 2004. Copping (2004) susținea că este singurul biopreparat care poate fi utilizat în combaterea pătării reticulare a orzului, fie ca tratament la semințe, fie ca tratament foliar. A urmat o perioadă în care produsul nu a mai primit aprobare, în special din cauza prezenței unui metabolit (DDR) cu proprietăți mutagene [EFSA, 2020].
În prezent, Pseudomonas chlororaphis MA 342 a primit aprobare pentru utilizare la cereale doar ca tratament la semințe. Aprobarea este datată 01.03.2023. Omologarea va expira în anul 2038 [după Official Journal of the European Union, 2023]. În România nu apare pe lista substanțelor omologate.

Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
CITEȘTE ȘI: În culturile de cereale apare făinarea
Un pericol pentru rapiță, Ceutorhynchus napi
Septorioza frunzelor, o boală care poate reduce producția de grâu
În culturile de cereale apare făinarea

Fungul Blumeria graminis produce boala numită „făinare”. Acest fung este prezent în culturile de cereale an de an, cu frecvențe și intensități diferite de atac, în funcție de condițiile climatice. În toamnele și iernile blânde, în culturile de cereale păioase sunt observate simptomele tipice ale patogenului. În articolul de față găsiți informații utile despre biologia patogenului, simptomatologia, pagubele produse și strategia de combatere.
În culturile de grâu și orz din vestul țării (Arad - Crișana), fungul Blumeria graminis a produs infecții în culturile de grâu și orz mai avansate în vegetație și cu densitate mai mare de plante/metru pătrat. Condițiile climatice înregistrate în prima și a doua decadă a lunii martie 2026 (vreme răcoroasă și umedă) au favorizat patogenia.
În culturile de grâu și orz verificate, am observat miceliile albe, bumbăcoase, pe tulpini și frunze. În cazul în care vremea răcoroasă și umedă se menține, patogenul va urca în etajele superioare ale plantelor, pe măsură ce acestea se dezvoltă. Temperaturile ridicate și lipsa precipitațiilor opresc evoluția făinării cerealelor păioase.
Pe lângă factorii climatici, infecțiile sunt susținute și de factorii tehnologici (monocultura sau absenţa rotaţiei, densitatea mare a plantelor, irigarea, excesul sau carenţa elementelor nutritive, întârzierea semănatului).

Recunoașterea simptomelor
Simptomele produse de Blumeria graminis la orz și grâu sunt foarte ușor de recunoscut, neputând fi confundate cu simptomele produse de alți patogeni foliari. Atacă toate organele aeriene ale plantelor (frunze, teci, tulpini, spice, ariste).
Tabloul simptomatic al bolii:
-
După realizarea infecției, pe frunzele bazale apar pete clorotice sau galbene;
-
La suprafața petelor de pe frunze, pe măsură ce patogenul evoluează, se formează aglomerări de micelii mici, albe și cu aspect pâslos. Miceliile albe pot fi izolate sau se pot uni;
-
În condiții favorabile, pete acoperite de micelii vor apărea și pe frunzele din etajele superioare, pe tecile frunzelor, pe tulpini (miceliile le cuprind de jur împrejur ca un manșon) și în final pe spice;
-
Miceliile albe de pe organele atacate își vor schimba culoarea (de la alb la gălbui) pe măsură ce boala evoluează, căpătând aspect prăfos, făinos. Este semn că ciuperca sporulează (se formează lanțurile de conidii sau oidii). Datorită aspectului făinos, boala a primit numele popular de „făinare”;
-
Pe măsură ce plantele devin mature, aglomerările de hife miceliene, devin gri şi apoi uşor brune la culoare. În această etapă, pot fi observate în micelii corpușoare mici, negre, asemănătoare cu boabele de piper (peritecii sau cleistotecii cu asce şi ascospori). Formarea cleistoteciilor reprezintă sporogeneza telomorfă sau sexuată a ciupercii sau „faza galben – roşcată”.
-
Sub pâsla miceliană, uşor desprinsă cu degetele mâinilor, ţesuturile plantelor sunt brune, necrotice sau moarte [Hatman et al., 1989; Eliade, 1990; Lipps, 1996; Baicu et Seşan, 1996; Popescu, 1998, 2005].
În condiții foarte favorabile, la soiurile sensibile și în zonele unde sunt prezente patotipuri cu virulență ridicată, manifestarea la exteriorul plantelor, specifică ciupercii Blumeria graminis, devine severă, amplă, adică ia caracter de masă sau de epidemie şi chiar de pandemie [Prescott et al., 1986; Popescu, 1998; Bissonette, 2002].
Supraviețuirea patogenului peste iarnă
Fungul ierneaza în anotimpul rece sub formă de cleistotecii pe samulastra de grâu și orz infectat. Pe lângă cleistotecii, patogenul poate ierna și sub formă de micelii pe plantele de grâu şi orz, putând produce conidii ce pot fi responsabile de infecțiile inițiale. Iernarea şi perpetuarea de la un an la altul a fost şi este studiată de diferiţi cercetători, dar ca şi alte probleme şi în aceasta sunt multe lucruri neelucidate sau controversate.
Făinare la grâu, 17.03.2026

Realizarea infecțiilor
Infecțiile cu Blumeria graminis pot apărea încă din toamnă dacă vremea permite. Uneori, în iernile blânde se pot observa micelii albe pe frunzele tinerelor plăntuțe. Infecţiile de toamnă constituie sursa principală de răspândire a bolii, miceliul rezistând peste iarnă [Hulea et al., 1975; Hatman et al., 1989; Popescu, 1998; Bissonnette, 2002].
Primăvara, primele infecții sunt produse de ascosporii eliberați din ascele aflate în cleistotecii cât și de conidiile produse de miceliile care iernează. Cleistoteciile se formează pe frunze, pe tulpini şi teci (iernează pe acestea), iar în primăvara următoare ascosporii eliberați produc infecţiile primare [Sandu-Ville, 1967; Eliade, 1990; Davis et al., 2002].
Infecțiile secundare în sezonul de vegetație sunt produse în mod repetat de conidiile care se formează la suprafața miceliilor când ciuperca sporulează (sporulare asexuată). Conidiile sunt purtate de vânt pentru ciclul secundar al bolii la intervale de 10 zile.
Făinare la orz, 17.03.2026

Condiții climatice favorabile infecțiilor
Factorii de mediu contează cel mai mult în realizarea infecțiilor.
Realizarea infecțiilor este în strânsă corelație cu următorii parametri climatici:
-
Temperatura. Fungul Blumeria graminis realizează infecţia cerealelor şi-şi manifestă patogenitatea în limite largi de temperatură. Cu toate acestea, ciuperca este virulentă în condiții de răcoare. Asta înseamnă că preferă temperaturile cuprinse între 17 - 220C [Prescott et al., 1986; Williams et Littlefield, 1995] sau 15 - 250C [Kochourek et Vechet, 1984; Bailey et al., 1995; Lipps, 1996]. Pe măsură ce temperaturile trec de 250C, patogenul nu mai infectează.
-
Umiditatea (roua, precipitațiile, umiditatea relativă a aerului). Umiditatea relativă a aerului şi precipitaţiile interferează pozitiv cu gradul de atac al ciupercii, dar cu o intensitate redusă la jumătate faţă de rouă. S-a constatat că ciuperca poate fi mai agresivă la valori mai scăzute ale umidității (37 - 56%) decât la o atmosferă cu hidroscopicitate de 79 - 97% (Sandu-Ville, 1967; Kocourek et Vechet, 1984; Eliade, 1990; Yang et al., 1992; Friedrich, 1995 a şi b; Deacon, 1997, 2006; Chet, 2003; Cotuna et Popescu, 2005b). Alți autori arată că făinarea poate fi puternic extensivă atunci când umiditatea relativă este cuprinsă între 85% și 100% (în prezența sau lipsa ploilor) - [Kochourek et Vechet, 1984; Prescott et al., 1986; Bailey et al., 1995; Williams et Littlefield, 1995; Lipps, 1996]. Ploile puternice nu sunt favorabile producerii de spori sau creşterii miceliului pe suprafaţa frunzelor [Evans, 1997; Chet, 2003].
-
Lumina. Însuşirile de patogenitate ale ciupercii sunt influenţate şi de lumină şi de întuneric. La întuneric lanţurile de oidii sunt mai lungi, au vitalitate scăzută şi o slabă putere de infecţiozitate datorită conţinutului scăzut de carbohidraţi [Sandu-Ville, 1967; Kocourek et Vechet, 1984; Eliade, 1990].
-
Nebulozitatea de 3 - 6 este la limita semnificaţiei [Deacon, 1997, 2006; Chet, 2003; Cotuna et Popescu, 2005b].
-
Viteza vântului este importantă în diseminarea patogenului în interiorul plantelor și la distanțe mai mari [Eliade, 1990; Cotuna et Popescu, 2005b].
Managementul integrat al făinării cerealelor
Făinarea cerealelor poate fi combătută prin utilizarea echilibrată a măsurilor profilactice, chimice și biologice. În România, de regulă patogenul nu pune probleme decât în anii extrem de favorabili infecțiilor și doar atunci când infecția ajunge la spic putem discuta de daune.

Măsuri profilactice
Aceste măsuri au rol important în prevenirea făinării la grâu dar și la alte cereale și constau în: respectarea rotaţiei culturilor; executarea corectă a lucrărilor solului; semănatul la date şi densităţi optime; folosirea soiurilor rezistente cu productivitate ridicată; utilizarea raţională a fertilizării; distrugerea samulastrei; irigaţia judicioasă acolo unde este cazul [Hatman et al., 1986; Iacob, 2003].
Măsurile de prevenție enumerate pot ține departe boala. Pe de altă parte, sunt și cele mai ieftine.
Măsuri chimice
Combaterea chimică trebuie să se facă la avertizare, după cum urmează:
-
După înfrățit, când pe ultimele trei frunze sunt peste 25 pete pâsloase;
-
Înainte de înflorit, când pe frunza stindard sunt peste 25 pete pâsloase (PED-ul sau pragul economic de dăunare) și factorii climatici (temperatură, umiditate, ploaie, ceaţă, rouă) continuă să se întrunească în limite optime pentru dezvoltarea bolii [Popescu, 1998].
De reținut că, stropirile aplicate la faza de un nod (stadiu de creştere GS 31) au controlat de timpuriu făinarea. De asemenea, cel mai bun control a fost asociat cu stropirile aplicate la emergerea frunzei stindard (GS 39 – 43) sau apariţia spicului (GS 59), stadii dezvoltate înainte de creşterea atacului. Stropirile aplicate în fenofazele amintite au determinat o bună protecţie a spicului [Harwick et al., 1994].
Fungicidele omologate în România pentru combaterea făinării cerealelor (dar și pentru alte boli ale cerealelor) sunt: Azoxistrobin; Azoxistrobin + protioconazol; Azoxistrobin + difenoconazol + tebuconazol; Azoxitrobin + tebuconazol; Azoxistrobin + fluxapyroxad; Protioconazol + tebuconazol; protioconazol + spiroxamină + trifloxistrobin; Protioconazol + spiroxamină + tebuconazol; Bixafen + spiroxamină + trifloxistrobin; Bixafen + protioconazol + spiroxamină; Bixafen + protioconazol; Bixafen + tebuconazol; Difenoconazol; Metrafenonă; Sulf; Ciprodinil; Ciflufenamid; Piriofenonă; Piraclostrobin; Fluxapyroxad; Fenpropidin; Fenpicoxamidă + protioconazol; Fenpropidin + protioconazol; Fluxapyroxad + mefentrifluconazol; Fluxapyroxad + metconazol; Fluxapyroxad + piraclostrobin; Mefentrifluconazol + piraclostrobin; Mefentrifluconazol + protioconazol; Mefentrifluconazol + metrafenonă + piraclostrobin; Mefentrifluconazol; Metconazol; Protioconazol; Protioconazol + spiroxamină; Protioconazol + trifloxistrobin; Tebuconazol; Tebuconazol + trifloxistrobin; Tetraconazol; Boscalid + kresoxim metil; Benzovindiflupir; Bromuconazol + tebuconazol; Difenoconazol + fluxapiroxad; Proquinazid; Proquinazid + protioconazol; Kresoxim - metil + mefentrifluconazol [după aplicația PESTICIDE 2.24.3.1, 2024].
Măsuri biologice
În culturile de cereale, măsurile biologice aproape că nu există. Având în vedere contextul actual (multe pesticide sunt retrase) există interes la nivel mondial pentru mai mulți agenți biologici care ar putea fi utilizați în combaterea făinării cerealelor. Aceștia sunt: Bacillus subtilis, B. chitinospora, B. pumilus, Pseudomonas fluorescens, Rhodotaula sp. (Xiaoxi & Wenhong, 2011; Shahin et al., 2019).

Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
CITEȘTE ȘI: Un pericol pentru rapiță, Ceutorhynchus napi
Septorioza frunzelor, o boală care poate reduce producția de grâu
Gândacul ghebos, dăunătorul care poate compromite culturile de cereale
Patogen nou la rapiță în România! Prima raportare a fungului Macrophomina phaseolina

Macrophomina phaseolina la floarea-soarelui am raportat-o pentru prima dată în anul 2021 în județul Timiș. De atunci, fungul face ravagii prin culturi. Motivele le știm cu toții. Iată că, a venit timpul să raportez acest fung și la rapiță.
În urma unui control fitosanitar făcut în județul Argeș împreună cu echipa OGOR al cărei partener sunt, am identificat acest patogen la o cultură de rapiță din zona Șerbănești. Cultura era deja recoltată. Am prelevat plante (tulpini rămase pe câmp) pentru a evalua atacul de Baris coerulescens. Nu mică mi-a fost mirarea când am găsit microscleroți de Macrophomina phaseolina în zona coletului și în rădăcini. Menționez că, fermierii din această zonă practică tehnologii no-till. De asemenea, rotațiile practicate sunt scurte. Ce înseamnă asta? Răspuns de la un fermier: „floarea soarelui - grâu, rapiță - porumb – floarea-soarelui etc”.

Vă atrag atenția că, pe termen lung vor apărea probleme foarte mari cu privire la patogenii și dăunătorii specifici. Mai mult, porumbul este și el plantă gazdă pentru Macrophomina phaseolina. De aceea, îmi voi îndrepta atenția și către porumb în acest an. E posibil să avem surprize.
Prezența acestui fung la rapiță a fost raportată în Statele Unite, Argentina, Australia (2004, 2006, 2009). Sunt puține lucrări pe subiect, cele mai multe studiind Macrophomina phaseolina la modul general (are mai multe plante gazdă, foarte multe).
Vom vedea în viitor care va fi impactul acestui patogen asupra producției de rapiță. La floarea-soarelui vedem deja.
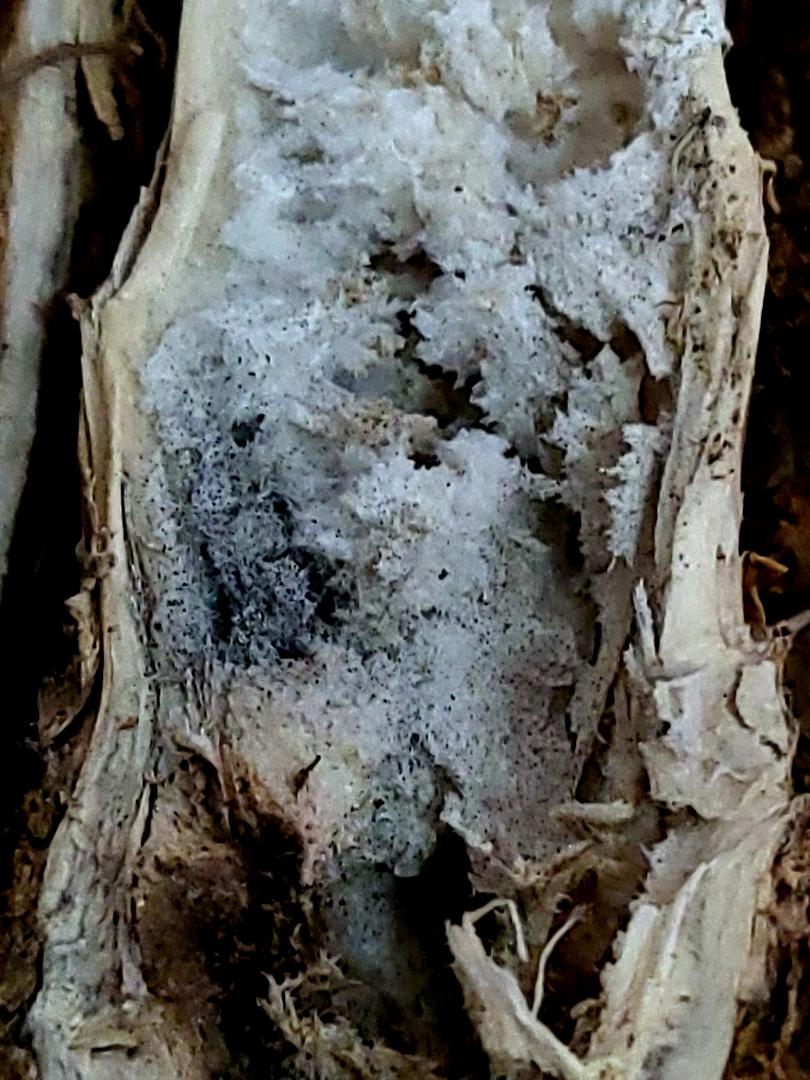
Cum se manifestă Macrophomina phaseolina la rapiță
Primele simptome pot fi observate în lanul de rapiță în preajma înfloritului. Plantele infectate prezintă simptom de ofilire generală atunci când temperaturile sunt ridicate în timpul amiezei (Schwartz & Gent, 2004). Spre seară plantele își revin (scad temperaturile). La final, ofilirea ia caracter permanent, iar plantele se usucă. De regulă, în lan se observă de la distanță vetre alungite (urmează direcția rândurilor) cu plante ofilite (Gaetán et al., 2006).

În timpul umplerii semințelor se observă cel mai bine plantele bolnave. Simptomele care apar sunt:
-
Ofilirea frunzelor;
-
Pete longitudinale de culoare maronie pe tulpina principală, cât și pe ramurile laterale;
-
Ramurile laterale se pot usca;
-
În cazurile grave pot apărea despicări longitudinale ale tulpinilor în zona bazală;
-
Senescența prematură;
-
Stagnarea în creștere a plantelor/aspect de pipernicire;
-
Silicve goale sau cu semințe șiștave;
-
Pe tulpini în zona bazală se formează pete de culoare gri - argintiu până la negru;
-
În măduvă, în rădăcinile pivotante și în țesutul vascular al bazei tulpinii se formează microscleroți foarte mici de culoare neagră care dau aspect cărbunos;
-
Plantele bolnave se smulg ușor din sol;
-
Moartea plantelor, mai ales în timpul perioadei de umplere a semințelor (Gaetán et al., 2006; Khangura & Aberra, 2009).
La infecții masive, plantele se vor usca în fenofaza de umplere a semințelor. Astfel de plante se vor smulge ușor din sol deoarece rădăcinile și partea bazală a tulpinii sunt distruse, putrede, cu aspect sfărâmicios (Gaetán et al., 2006; Khangura & Aberra, 2009).

Realizarea infecțiilor
Infecțiile se realizează în primele etape de viață ale rapiței în condiții de temperaturi ridicate și sol uscat. După Gaetán et al. (2006), Macrophomina phaseolina este deosebit de dăunătoare atunci când temperatura solului este de 320C și plantele sunt stresate de secetă. Microscleroții rezistă în sol mai mult de zece ani.
Putem combate acest patogen?
Metodele profilactice sunt cele mai importante. Se recomandă rotația la patru ani sau mai mult, cultivarea hibrizilor rezistenți sau toleranți, irigarea în perioadele secetoase și calde, controlul buruienilor.
Măsurile chimice nu dau rezultatele scontate. De asemenea, nu există fungicide omologate pentru acest patogen (Schwartz & Gent, 2004).
BibliografieGaetán S. A., Fernandez L., Madia M., 2006. Occurrence of Charcoal Rot Caused by Macrophomina phaseolina on Canola in Argentina, Plant Disease, 90 (4): 524 - 524.Khangura R., Aberra M., 2009. First Report of Charcoal Rot on Canola by Macrophomina Phaseolina in Western Australia, Plant Disease, 2009, 93:6, 666 - 666.Schwartz H. F., Gent D. H., 2004. Canola and Mustard - Charcoal Rot, High Plains IPM Guide, a cooperative effort of the University of Wyoming, University of Nebraska, Colorado State University anf Montana State University 2 pp., disponibil pe bugwoodcloud.org/bugwoodwiki/CharcoalRot - CanolaMustard.pdf
Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
Consecințele atacului gărgărițelor tulpinilor de rapiță

În unele sole de rapiță din Timiș plantele au căzut în vetre, frângându-se sub acțiunea vânturilor puternice și a ploilor abundente. În urma observațiilor făcute, am constatat că plantele erau pline de orificii produse de larvele gărgărițelor tulpinilor. Pe zonele lezate erau pete specifice fungului Phoma lingam cu picnidii negricioase evidente.
Plante frânte din cauza gărgărițelor tulpinilor și fungului Phoma lingam

Gărgărițele și fungul Phoma lingam au determinat putrezirea țesuturilor, iar plantele s-au rupt foarte ușor. Cu siguranță, fermierul a ratat momentele optime de combatere gândindu-se că activitatea gărgărițelor a fost oprită de vremea rece. Din păcate nu a fost așa, gărgărițele și-au continuat dezvoltarea nestingherite.
Vatră cu plante frânte. Silicvele au început să se îngălbenească, plantele se vor usca la final

Cunoaștem că gărgărițele tulpinilor au importanță economică deoarece pot distruge plantele prin reducerea creșterii lor, afectând în final producția de semințe. În principal, stadiul larvar este cel care produce daune rapiței, dar și altor brassicaceae (varză, conopidă, ridichi, muștar). Din cauza atacului larvelor, tulpinile se deformează, crapă și se pot frânge foarte ușor. Zonele lezate sunt o poartă de intrare pentru agenții patogeni ai tulpinii, dar și pentru alți fungi ce pot produce putrezirea. Uneori plantele ramifică excesiv, nu mai cresc și în cazurile grave chiar nu mai formează silicve [Roșca et al., 2011]. Pagubele pot ajunge uneori la 50% din producție și chiar mai mult.
Phoma lingam și gărgărițe

Phoma lingam pe tulpini

Larvele s-au retras din tulpini

Orificii produse de larve

Ce trebuie să facă un fermier care se confruntă cu o infestare masivă care i-a adus pagube?
Ar trebui să izoleze cultura, să îndepărteze resturile de plante care rămân după recoltare (sunt sursă de patogeni pentru anul următor) și să facă o arătură adâncă după recoltare (mai ales atunci când infestarea a fost mare deoarece larvele se împupează în sol).
Dacă arătura se execută mai târziu, va fi ineficientă, deoarece adulții de Ceutorhynchus pallidactylus iernează în afara culturii infestate. NU la fel se întâmplă cu Ceutorhynchus napi care rămâne în sol până în toamnă.
Se observă fructificațiile fungului Phoma lingam și albirea specifică

Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
CITEȘTE ȘI: Phoma lingam, atac la tulpinile de rapiță
Monitorizarea gărgărițelor tulpinilor de rapiță
Phoma lingam, atac la tulpinile de rapiță

În culturile de rapiță din Timiș este evident atacul de Phoma lingam la tulpini. Condițiile climatice de la sfârșitul acestei primăveri sunt propice infecțiilor. Simptomele pot fi observate mai ales la plantele atacate de gărgărițele Ceutorhynchus napi și pallidactylus.
Înnegrirea tulpinii este o boală cu importanță economică a rapiței, în Europa fiind raportate periodic epidemii. Se cunoaște că, în culturile de rapiță pot coexista două specii de Leptosphaeria: Leptosphaeria maculans (Desm.) Ces. & Not. (forma sexuată) cu forma asexuată Phoma lingam (Tode ex. Schw.) Desm. (mai agresivă) și Leptosphaeria biglobosa (mai puțin agresivă).
Phoma lingam, atac la tulpini. 20 mai 2025, Bucovăț - Timiș

Phoma lingam este un fung care, în condiții climatice favorabile poate produce pagube importante în producție. Majoritatea simptomelor produse de Leptosphaeria maculans se instalează după o perioadă de umectare a frunzelor cuprinsă între 4 - 48 de ore și temperaturi de 12 - 200C. Condițiile climatice sunt foarte importante în evoluția patogeniei. Astfel, infecțiile se pot instala după cinci zile la temperaturi de 200C și după două săptămâni la 80C [Biddulph et al., 1999b]
Simptomele produse de Phoma lingam sunt foarte ușor de recunoscut. Patogenul poate ataca plantele pe tot parcursul perioadei de vegetație, începând cu faza de cotiledon. Cel mai adesea frunzele sunt primele infectate. Atunci când vremea este prielnică fungului, acesta trece pe tulpini și chiar pe silicve (la infecții masive).
Se observă la suprafața petei picnidiile negricioase ale fungului Phoma lingam

Pentru că în această perioadă sunt prezente infecțiile la tulpini readuc în atenție simptomele specifice. Această formă de atac este cea mai periculoasă pentru rapiță.
Petele de pe tulpinile verzi pot avea culoare brună și chiar negricioasă. În condiții favorabile, petele se transformă în leziuni deschise ce pot predispune la putrezire.

Infecția din zona bazei tulpinilor este considerată cea mai periculoasă formă de atac. În prima fază, leziunile de la baza tulpinii au culoare brună deschis după care se înnegresc (nu întodeauna) și înconjoară tulpina (aspect de necroză). Uneori, petele de pe tulpini sunt alungite, au culoare cenușie și marginea de culoare brună închis (observație personală). Ca și la frunze, la suprafața leziunilor înnegrite pot fi observate picnidiile fungului ca niște bobițe foarte mici de culoare neagră. În condiții de umiditate, picnidiile expulzează mase gelatinoase de culoare roz deschis pline de conidii. În condiții de uscăciune, acestea iau diferite forme sinuoase, cel mai adesea de cârcei. Pot fi observate chiar cu ochiul liber sau cu o lupă de mână.
În prezența umidității, picnidiile expulzează mase gelatinoase de culoare roz deschis pline de conidii care vor realiza noi infecții. Foto la lupa binocular. Probă din rapiță - Bucovăț (Timiș)

Înnegrirea bazei tulpinii este periculoasă în timpul înfloritului și al formării silicvelor și maturizării semințelor. Petele de la baza tulpinii se pot uni, după care se rup formând putregaiuri uscate (leziuni deschise care putrezesc). Această formă de manifestare este considerată cea mai dăunătoare [Paul și Rawlinson, 1992].
Terminologia utilizată în fitopatologie pentru descrierea acestui simptom include mai mulți termeni:
-
înnegrirea bazei tulpinii (blackleg în engleză);
-
leziuni necrotice deschise (basal canker în engleză);
-
necroze cu aspect coronat care înconjoară baza tulpinii (crown canker);
-
necroze cu aspect de guler (collar rot) etc.
Infecția la silicve se manifestă sub forma unor leziuni deschise de culoare brună - deschis și margini închise la culoare care fac delimitarea față de țesutul sănătos.
Managementul integrat al fungului Leptosphaeria maculans
Leptosphaeria maculans poate produce pagube importante culturilor de rapiță în condiții de vreme umedă, când există pericolul de atac al bazei tulpinilor. Strategiile de management ale acestui fung, trebuie elaborate cu atenție, în așa fel încât accentul să cadă pe măsurile de prevenție și mai puțin pe cele chimice. Nu sunt excluse nici măsurile biologice (dacă există biopreparate).
Conidii de Phoma lingam la microscop

Măsuri profilactice
Acest set de măsuri este esențial în controlul bolii „înnegrirea tulpinilor”. Se recomandă respectarea următoarelor măsuri:
-
Cultivarea hibrizilor rezistenți poate aduce economii în bugetul fermierilor;
-
Utilizarea la semănat a semințelor libere de patogeni, certificate;
-
Rotația la patru ani scade foarte mult riscul infecțiilor, pe când rotațiile scurte cresc riscul de infecție. Aici sunt multe probleme;
-
Distrugerea resturilor vegetale după recoltare și îngroparea lor - patogenul supraviețuiește pe reziduurile vegetale, nu în sol, timp de 2 - 3 ani;
-
Înființarea noilor culturi la o distanță mai mare de 500 m față de vechea cultură de rapiță.
În anii favorabili infecțiilor, respectarea măsurilor profilactice vă poate proteja de infecțiile cu Leptosphaeria maculans.
Măsuri chimice
Este important ca sămânța să fie tratată înainte de semănat, deoarece patogenul poate supraviețui în semințe. Pentru tratarea semințelor, în România este omologat un singur produs: Fluopicolid + fluoxastrobin.
În perioada de vegetație, tratamentele se execută ținând cont de biologia patogenului, de condițiile climatice prognozate și de fenofaza plantelor.
Un prim tratament poate fi executat în fenofaza de 2 - 6 frunze sau 4 - 8 (toamna). Acest tratament are rolul de a scădea riscul infecțiilor ulterioare. Scopul tratamentelor foliare este de reduce riscul apariției infecțiilor la tulpină care sunt și cele mai periculoase. În acest sens, trebuie monitorizați factorii climatici temperatură și precipitații. Dacă se întrunesc mai multe zile cu precipitații după însămânțat este bine să luăm în calcul executarea unui tratament. De asemenea, ar trebui urmărită emisia ascosporilor. Această lucrare ar trebui făcută de către specialiști (din păcate nu se face). Prima depistare a ascosporilor (mai mult de 20 ascospori/zi) coroborată cu condițiile climatice, sensibilitatea hibridului, stadiul de creștere și vigoarea plantelor sunt factori ce vă pot ajuta în luarea unei decizii corecte cu privire la momentul optim de efectuare a tratamentului chimic.
Atac de larve de Ceutorhynchus napi și/sau pallidactylus. Tulpina este atacată și de fungul Phoma lingam

Al doilea tratament în vegetație (primăvara) se face cu respectarea criteriilor menționate mai sus. Primăvara, emisiile maxime de ascospori au loc de obicei în luna aprilie. Observațiile efectuate de mine în Banat, arată că, în unele primăveri, emisiile de ascospori pot apărea încă de la începutul lunii martie.
În cazul Phomei lingam, specialiștii apreciază că, de fapt nu există „momente optime” stabilite pentru combaterea acestui patogen, de aceea este important să monitorizăm patogenul pe întreaga perioadă de vegetație și să intervenim când situația din teren impune asta.
În România sunt omologate pentru tratamentele în vegetație următoarele fungicide: Azoxistrobin; Azoxistrobin + difenoconazol; Mepiquat clorură + piraclostrobin + prohexadion de calciu; Metconazol; Mepiquat clorură + metconazol; Tebuconazol; Difenoconazol; Protioconazol; Boscalid; Difenoconazol + protioconazol; Difenoconazol + tebuconazol; Difenoconazol + paclobutrazol [după APLICAȚIA PESTICIDE 2.25.4.1, 2025].
Măsuri biologice
În România sunt omologate două biopreparate: agentul biologic M1 x 106 oospores/g Pythium oligandrum pentru tratamente în vegetație; agentul biologic Bacillus amyloliquefaciens, tulpina MB 1600 (controlează Leptosphaeria maculans), tratament la sămânță.
Interesul pentru utilizarea agenților biologici este în creștere. Leptosphaeria maculans ar putea fi controlată cu ajutorul bacteriei Paenibacillus polymyxa PKB1 care produce peptide ce pot inhiba sau ține sub control infecțiile la frunze și tulpini [Kharbanda et al., 1999]. Multe studii există cu privire la utilizarea acestei bacterii la rapiță.
BibliografieBiddulph J. E., Fitt B. D. L., Leech P. K., Welham S. J., Gladders P., 1999b - Effects of temperature and wetness duration on infection of oilseed rape by ascospores of Leptosphaeria maculans (stem canker). European Journal of Plant Pathology 105, 769 - 81.Kharbanda P. D., Yang J., Beatty P., Jensen S., Tewari J. P., 1999 - Biocontrol of Leptosphaeria maculans and other pathogens of canola with Paenibacillus polymyxa PKB1. Proceedings of the 10th International Rapeseed Congress, 1999. Canberra, Australia.Paul V. H., Rawlinson C. J., 1992 - Diseases and Pests of Rape. Gelsenkirchen-Buer, Germany: Verlag Th. Mann.
Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
CITEȘTE ȘI: Monitorizarea dăunătorilor porumbului
De ce se albesc spicele de grâu?
De ce se albesc spicele de grâu?

Am parcurs o primăvară capricioasă, umedă și răcoroasă care a influențat dezvoltarea plantelor cultivate. În această perioadă, prin unele zone există culturi de grâu care prezintă simptome de albire și sterilitate distală. De ce se albesc spicele de grâu? Cauzele albirii sunt multe, de aceea simptomele trebuie atent analizate pentru un diagnostic corect.
Albire și sterilitate distală

În cele ce urmează voi enumera principalele cauze ale albirii spicelor.
Albire produsă de atacul ploșnițelor cerealelor (Eurygaster integriceps, E. maura, E. austriaca, Aelia acuminata, Aelia rostrata).
Organele atacate de ploșnițe sunt: tulpina, frunzele, spicul și cariopsele. Adulții hibernanți se hrănesc pe organele vegetative. La locul înțepăturii, apare o mică umflătură (con salivar) înconjurată de o zonă decolorată, gălbuie. Frunzele atacate, se îngălbenesc, se răsucesc și se usucă de la locul unde ploșnița a înțepat, atârnând ca un fir de ață mai gros. Din cauza atacului, uneori spicele rămân în burduf. Dacă ies din burduf, pot avea aristele ondulate (la soiurile aristate) sau poate apărea fenomen de sterilitate parțială sau totală și chiar albirea vârfului în situațiile grave.
Ploșnițe la data de 21 mai 2025, Bucovăț - Timiș

Atacul produs de adulții hibernanți produce de regulă pierderi cantitative, nesemnificative. Periculos este atacul larvelor la spic care duce la pierderi calitative foarte periculoase, cum ar fi degradarea glutenului sub acțiunea enzimelor secretate de ploșnițe [Rajabi, 2000]. Pierderea elasticității glutenului duce la deprecierea calităților de panificație. Cariopsele atacate se recunosc ușor datorită înțepăturilor cu aspect de punct negricios înconjurat de o zonă de decolorare. Uneori punctul negricios nu este evident. Alteori, cariopsele atacate sunt zbârcite. Este bine ca procentul de boabe înțepate să nu treacă de 2%. Dacă trece de acest procent, calitatea pentru panificație a grâului începe să scadă [Rajabi, 2000]. După Roșca et al. (2011), la 15 - 20% boabe atacate, grâul nu mai poate merge către panificație.
Atac ploșniță

Albire produsă de musca de Hessa - generația de primăvară. Plantele atacate de larvele acestei generații se recunosc după îndoirea care apare la ultimul sau penultimul nod. În aceste zone paiul se albește comparativ cu restul tulpinii care rămâne verde. Uneori spicul are port erect, iar florile pot avorta. Astfel de simptome se pot datora și altor dăunători, dar și unor fungi. Este important să verificăm zona afectată pentru a exclude fungii sau alți dăunători [Popov, 1999 citat de Roșca et al., 2011].
Albire produsă de atacul tripșilor (larvele sunt mai periculoase). În urma hrănirii apare simptomul de spic albit parțial sau total, sterilitate parțială și umplerea incompletă a cariopselor.

Albire produsă de Fusarium graminearum. Forma cea mai gravă de atac este după înspicare. Spicele, iniţial se albesc parţial (câteva spiculeţe) sau total, apoi se înroşesc şi se acoperă cu un înveliş micelian, alb – roz sau alb – rubiniu, uneori portocaliu - somon, pe care se observă sporodochiile ciupercii (forma imperfectă). Pe spicele înroşite (pe palee, ariste sau boabe) se observă puncte negre care sunt periteciile ciupercii (forma perfectă). Cariopsele infectate sau fuzariate rămân mici, zbârcite, cenuşii sau rozii iar germinaţia şi puterea de străbatere va fi slabă [Popescu, 2005].
Albire produsă de înghețuri în timpul fenofazelor de burduf, înspicare și înflorire. Brumele și temperaturile scăzute sunt și ele periculoase. Grâul este sensibil la temperaturi de 00C, minus 10C, minus 20C ce survin în fenofazele amintite. Pe lângă asta, poate suferi chiar și la temperaturi de până la 50C, atunci când acestea se întind pe mai multe zile consecutiv.
Lan de grâu albit în comparație cu lan de grâu vecin care nu are astfel de simptome. 21 mai 2025, Arad

Înghețurile sau temperaturile scăzute pot produce daune severe. Un îngheț sever în timpul înfloritului va duce la sterilitate și implicit pagube în producție. Atunci când înghețul apare în fenofaza de burduf (este suficientă temperatura de minus 20C), putem observa următoarele simptome: vârful spicului rămâne captiv în burduf, iar baza spicului iese, brunificări ale tulpinii, decolorarea frunzelor, sterilitate. Dacă spicele ies normal din burduf, vor rămâne galbene sau chiar albe. Când vedem un astfel de simptom trebuie să știm că spicul este compromis.
Dacă în timpul înspicatului, intervin temperaturi de minus 10C, putem observa ariste albite. Înghețurile care albesc aristele, de regulă vor afecta și părțile mascule ale florii. De asemenea, mai putem observa pe tulpina de sub spic apariția unei zone de țesut clorotic (inel de îngheț). În această zonă, spicele se pot rupe, mai ales în condiții de vânt.
Cele mai periculoase sunt înghețurile ce apar în timpul înfloritului (minus 10C), grâul fiind extrem de sensibil în această perioadă. Expunerea la îngheț ucide organele florale mascule și provoacă sterilitate. Sterilitatea poate fi totală, completă sau parțială. Spiculețele pot fi goale sau parțial umplute. De asemenea, aristele se albesc.

Albirea prematură a spicelor produsă de fungii Rhizoctonia cerealis, Gaeumannomyces graminis, Pseudocercosporella herpotrichoides.
Lan de grâu cu spice albite distal. Ipoteze: îngheț la burduf, temperaturi scăzute, greșeli în tehnologie (tratamente fungicide, erbicide, aminoacizi aplicate la temperaturi scăzute). Nu ne putem pronunța încă dar analizăm.

Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
CITEȘTE ȘI: Plutella xylostella - molia verzei, prezentă în culturile de rapiță
Infecții produse de fungul Rhizoctonia cerealis
În culturile de cereale au apărut primele infecții produse de fungul Rhizoctonia cerealis

Condițiile climatice din această primăvară sunt propice fungilor care se dezvoltă în condiții de vreme răcoroasă și umedă. Aceștia sunt: Rhizoctonia cerealis (rizoctonioza), Pseudocercosporella herpotrichoides (pătarea în ochi a bazei tulpinii), Gaummannomyces graminis (îngenuncherea cerealelor păioase), Puccinia striiformis (rugina galbenă). Frigul a încetinit activitatea fungilor foliari, aceștia fiind prezenți în culturile de cereale, fără a pune probleme deosebite. Prin culturile din Banat am observat la această dată simptome produse de fungii Septoria tritici, Drechslera tritici repentis, Pyrenophora teres, Blumeria graminis. În funcție de zonele climatice, starea fitosanitară a culturilor poate fi diferită. Acolo unde a plouat, Septoria tritici este prezentă cu intensitate de atac mai ridicată.
Vremea umedă și răcoroasă a favorizat dezvoltarea fungului Rhizoctonia spp., iar primele infecții au apărut în culturile de grâu verificate. De aceea vă îndrum să începeți verificarea culturilor. Controlați plantele în zona bazală pentru a descoperi simptomele. Dacă vremea va fi favorabilă acestui patogen, daunele vor apărea în timp scurt (vetre căzute). Vă reamintesc că anul trecut am avut astfel de situații în Banat și la grâu și la orz. Pagubele în producție au fost evidente în solele bolnave.
Pentru a gestiona acest patogen, Rhizoctonia spp., readuc în atenția dumneavoastră aspecte despre biologia, epidemiologia și combaterea rizoctoniozei.
Importanța economică și recunoașterea simptomelor
Rizoctonioza este o boală a bazei tulpinii cerealelor păioase ce apare frecvent în regiunile temperate [Lemanczyk, 2010]. După Cromey et al. (2002), producțiile putând fi afectate în procent de 5% în cazul infecțiilor moderate și până la 26% când infecțiile sunt severe. Cerealele de primăvară sunt mai predispuse la infecția cu Rhizoctonia cerealis decât cele de toamnă. Orzul este mai atacat decât grâul [Smiley et al., 2012; Lemańczyk et Kwaśna, 2013]. În Banat, boala apare în anii favorabili, cu frecvențe mai mari de atac la orz și mai mici la grâu.

Simptomele bolii pot apărea încă din toamnă dacă sunt condiții favorabile. Sunt atacate rădăcinile, tulpinile și tecile frunzelor.
Principalele simptome produse sunt:
-
Inițial, pe rădăcini apar pete de culoare deschisă care mai târziu capătă culoare brună. Țesuturile necrozate se rup cu ușurință lăsând descoperit cilindrul central. După Popescu (2005), tinerele plante au tendința de a forma alte rădăcini. Astfel, o plantă bolnavă va avea rădăcini mai multe dar scurte;
-
Pe tulpini și teci apar pete de formă ovală, alungite. Culoarea petelor este albicioasă – cenușie, iar de jur împrejur se formează o margine de culoare brun - închis. De multe ori aceste pete pot fi confundate cu ușurință cu cele produse de Pseudocercosporella herpotrichoides și Fusarium sp. [Murray et al., 2009]. Adesea, leziunile produse de patogen sunt superficiale;
-
Rareori, în condiții de vreme rece și umiditate mare în zona bazei tulpinilor, miceliul ciupercii străpunge teaca și infectează tulpina. Enzimele secretate produc necroza țesuturilor, căderea plantelor sub acțiunea ploilor și vântului, albirea prematură a spicelor, șiștăvirea cariopselor [Cromey et al., 2005];
-
Toamnele și primăverile reci sunt favorabile infecțiilor cu Rhizoctonia cerealis.
Supraviețuirea patogenului
Peste anotimpul de iarnă, fungul supraviețuiește sub formă de miceliu sau scleroți (pseudoscleroți) în sol și materialul vegetal infectat. Acest patogen nu formează spori asexuați, iar stadiul telomorf este foarte rar întâlnit în natură [Carling et Sumner, 1992; Popescu, 2005; Lemańczyk et Kwaśna, 2013].
Supraviețuirea patogenului pe resturile vegetale constituie o sursă importantă de inocul pentru culturile de cereale, care în condiții climatice favorabile patogeniei pot fi infectate [Sneh et al., 1996].
Plantele bolnave au tendința să dezvolte noi rădăcini, mai multe și mai scurte. Sunt vizibile la suprafața solului

Realizarea infecțiilor, răspândire
Rhizoctonia este o ciupercă de sol cu creștere vegetativă mai mult sau mai puțin continuă, sub formă de miceliu ramificat, de culoare albă sau brună. Ciuperca se dezvoltă bine în condiții de sol cald și umed. Hifele miceliene cresc în sol și în materialul vegetal mort. În solurile uscate ciuperca devine latentă, formând un miceliu gros, cu pereți îngroșați sau produce microscleroți (cu diametrul de cel puțin 0,5 - 1,0 mm, uneori până la 5 mm), care îi permit să supraviețuiască timp de mulți ani, chiar în lipsa unei gazde [Papavizas et al., 1975]. Scleroții și/sau miceliul din sol sau de pe resturile vegetale descompuse infectează cerealele prin intermediul hifelor care pătrund în rădăcini, tulpini și teci [Naito, 2006].
Când se realizează infecția, hifele fungice cresc la suprafața organului atacat, formând manșoane miceliene care înconjoară de jur împrejur și permit ciupercii să pătrundă în țesuturile plantelor și să preia substanțele nutritive pentru creșterea și dezvoltarea fungică continuă. Ciuperca este atrasă de stimulenții chimici eliberați de celulele vegetale în creștere activă și/sau reziduurile vegetale descompuse. Pe măsură ce procesul de atracție continuă, hifa fungică va intra în contact cu planta și se va atașa de suprafața sa externă. După atașare, ciuperca continuă să crească pe suprafața externă a plantei și va provoca boala prin producerea unei structuri specializate de infecție (appresorium) care pătrunde în celula plantei.
Procesul de infecție este susținut de producerea multor enzime extracelulare diferite care degradează diferite componente ale pereților celulari ai plantelor (de exemplu, celuloză, cutină și pectină). Pe măsură ce ciuperca ucide celulele plantei, hifele continuă să crească și să colonizeze țesutul mort, formând adesea scleroți. Inoculul nou este produs pe sau în țesutul gazdă și un nou ciclu se repetă atunci când sunt disponibile noi substraturi [Naito, 2006].
După Popescu (2005), spațiul dintre teacă și tulpină reprezintă o zonă în care miceliul ciupercii se poate dezvolta foarte bine iar prin enzimele secretate produce necroza țesuturilor tulpinii în condiții de umiditate ridicată și vreme răcoroasă.
Rhizoctonia nu produce conidii (spori fungici asexuali) și rareori produce bazidiospori (spori sexuali). Cu toate acestea, de multe ori boala este produsă prin intermediul basidiosporilor.
Factorii care concură la instalarea infecțiilor la cereale sunt: virulența agentului patogen, sensibilitatea gazdei și condițiile favorabile de mediu [Francl, 2001].
Factorii climatici care susțin infecțiile:
-
Temperatura și umiditatea joacă un rol important în realizarea infecțiilor și supraviețuirea patogenului [Otten et al., 2004]. Temperatura din zona rădăcinilor este importantă în realizarea infecțiilor. La temperaturi cuprinse între 6 - 190C sau când există variații de la 16 la 270C pot apărea pagube semnificative [Gill et al., 2001]. Wiese (1987) arată că Rhizoctonia la cereale este favorizată îndeosebi de temperaturile cuprinse între 9 - 100C;
-
După Ogoshi (1996), tinerele plante de grâu pot fi atacate în condiții de vreme răcoroasă de izolate de Rhizoctonia care se dezvoltă în astfel de condiții;
-
Alte izolate ale fungului preferă vremea mai caldă;
-
Optimul termic pentru dezvoltarea ciupercii are loc în intervalul 16 - 200C [Sneh et al., 1996]. Canicula și lipsa umidității opresc subit patogenia [Popescu, 2005];
-
Boala este favorizată de resturile vegetale infectate din anul anterior, de solurile acide, densitatea mare a plantelor, solurile slab drenate, irigarea excesivă, umiditatea ridicată în zona bazei tulpinii asociată cu temperaturi ale solului între 17 - 230C [Pitt, 1966; Popescu, 2005].
Răspândirea ciupercii pe distanțe mari se realizează prin intermediul sporilor sexuați (basidiosporii) în condiții de umiditate ridicată (peste 90%) și vreme caldă (temperaturi mai mari sau egale cu 200C) - Naito, 1996; Agrios, 2005. Basidiosporii ajunși pe frunze pot produce infecții foliare. Rolul acestor spori nu este încă cunoscut în totalitate [Naito, 2006].

Combaterea integrată a rizoctoniozei cerealelor
Odată instalat, patogenul este greu de combătut. Adesea el este observat când este prea târziu și plantele sunt căzute la sol. Cel mai ușor poate fi identificat atacul la stadiul de creștere GS 39 (frunza stindard vizibilă) - Zadocks et al., 1974.
Măsurile profilactice
Profilaxia este deosebit de importantă în managementul acestei boli. Amintesc: utilizarea soiurilor rezistente; eliminarea sursei de inocul (resturi vegetale); eliminarea gazdelor voluntare; rotații corecte; efectuarea lucrărilor solului (arături sau măcar lucrarea solului până la adâncimea de 8 - 10 cm) - Cook et al., 2002; Fletcher et al., 2010.
De reținut că, rotația, lucrările solului și data semănatului influențează instalarea patogenului [Colbach et al., 1997].
La cerealele cultivate în sistem „no tillage” sau „minimum tillage”, rizoctonioza se poate instala, deoarece solul este bogat în materii organice pe care ciuperca supraviețuiște [Oros et al., 2013].
Măsurile chimice
Tratamentele chimice au eficacitate variabilă, între 0 și 80%. De regulă, tratamentul semințelor și tratamentele aplicate la cereale în perioada de vegetație ar trebui să protejeze plantele de infecție [Popescu, 2005].
Dintre fungicidele utilizate în combaterea patogenilor cerealelor păioase, fludioxonilul este menționat în unele studii ca fiind eficient în combaterea rizoctoniozei [Smiley et al., 2012].
În România sunt omologate următoarele substanțe doar pentru tratarea semințelor:
fludioxonil cu sedaxan - Sedaxanul este o substanță din grupa „pirazolecarboxamide” cu spectru larg de acțiune, recomandată pentru controlul rizoctoniozelor. În combinații cu alte fungicide, rezultatele sunt mai bune; Difenoconazol + fludioxonil + sedaxan [după Aplicația PESTICIDE 2.25.2.2, 2025].
Controlul biologic
De interes sunt bacteriile din genul Pseudomonas și fungii antagoniști din genul Trichoderma [Budge et al., 2009].
BibliografieAgrios G. N., 2005 - Plant Pathology. 3rd Ed. London, Elsevier Academic Press, 952 p.Budge G., Shaw M., Colyer A., Pietravalle S., Boonham N., 2009 - Molecular Tools to investigate Rhizoctonia solani Distribution in Soil. Plant Pathology, 58, 1071 - 1080Carling D. E., Sumner D. R., 1992 - Rhizoctonia. In L. L. Singleton, J. D. Mihail, C. M. Rush (Eds.), Methods for research on soilborne phytopathogenic fungi. St Paul: American Phytopathological Society Press, 157 - 165.Colbach N., Lucas P., Cavelier N., Cavelier A., 1997 - Influence of Cropping System On Sharp Eyespot in Winter Wheat. Crop Protection, 16, 415 - 422.Cook R. J., Schillinger W. F., Christensen N. W., 2002 - Rhizoctonia Root Rot and Take-All of Wheat in Diverse Direct-Seed Spring Cropping Systems. Canadian Journal of Plant Pathology, 24, 349 - 358.Cromey M. G., Butler R. C., Boddington H. J., Moorhead A. R., 2002 - Effects of sharp eyespot on yield of wheat (Triticum aestivum) in New Zealand. N Z J Crop Hort. 30(1): 9 – 17.Cromey M. G., Butler R. C., Munro C. A., Shorter S. C., 2005 - Susceptibility of New Zealand wheat cultivars to sharp eyespot. N Z Plant Prot. 58: 268 – 272.Fletcher J., Luster D., Bostock R., Burans J., Cardwell K., Gottwald T., Mcdaniel L., Royer M., Smith K., 2010 - Emerging infectious Plant Diseases. Emerging infectious Diseases (Scheld Wm Et Al. Eds.) Pp, 33 - 366.Francl L. J., 2001 - The Disease Triangle: A Plant Pathological Paradigm Revisited. Plant Health instructor Doi, 10.Gill J., Sivasithamparam K., Smettem K., 2001a - Effect of Soil Moisture At Different Temperatures On Rhizoctonia Root Rot of Wheat Seedlings. Plant and Soil, 231, 91 - 96.Henegar Monica et al., 2019 - Codexul produselor de protecție a plantelor omologate pentru utilizare în România, Editura Agroprint, Timișoara, 619 p.Lemanczyk G., 2010 - Occurrence of sharp eyespot in spring cereals grown in some regions of Poland. J. Plant. Prot. Res. 50.(4): 505 – 512.Lemanczyk G., Kwasna H., 2013 - Effects of sharp eyespot (Rhizoctonia cerealis) on yield and grain quality of winter wheat. Eur. J. Plant. Pathol. 135:187 – 200.Murray T. D., Parry D. W., Cattlin N. D., 2009 - Diseases of small grain cereal crops, Manson Publishing Ltd, 142 p.Naito S., 1996 - Basidiospore dispersal and survival. In: Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control. B. Sneh, S. Jabaji-Hare, S. Neate, and G. Dijst, eds. Kluwer Academic Publishers, Dordrecht, The Netherlands, 197 - 205.Naito S., 2006 - Ecological Studies On Teleomorphic and Anamorphic Stages in Rhizoctonia Fungi. Journal of General Plant Pathology, 72, 400 - 403.Ogoshi A., 1996 - Introduction - the Genus Rhizoctonia. in: Sneh B., Jabaji-Hare S., Neate S., Dijst G. (Eds.) Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control. Springer Netherlands, 1 - 9.Oros G., Naár Z., Magyar D., 2013 - Susceptibility of Wheat Varieties to Soil-Borne Rhizoctonia infection. American Journal of Plant Sciences, 4, 22 - 40.Otten W., Harris K., Young I. M., Ritz K., Gilligan C., 2004 - Preferential Spread of the Pathogenic FungusRhizoctonia solani Through Structured Soil. Soil Biology and Biochemistry, 36, 203 - 210.Papavizas G., Adams P., Lumsden R., Lewis J., Dow R., Ayers W., Kantzes J., 1975 - Ecology and Epidemiology of Rhizoctonia solani in Field Soil. Phytopathology, 65, 871 - 877.Pitt D., 1966 - Studies On Sharp Eyespot Disease of Cereals: Effects of the Disease On the Wheat Host and the incidence of Disease in the Field. Annals of Applied Biology, 58, 299 - 308.Popescu Gheorghe, 2005 - Tratat de patologia plantelor, vol. II, Ed. Eurobit, 341 p.Smiley R., Paulitz T., Marshal J., 2012 - Controling Root and Crown Diseases of Small Grain Cereales, PNW 639, 9 p., accesat la data de 20.12.2020.Sneh B., Jabaji - Hare S., Neate S., Dijst G., 1996 - Rhizoctonia species: Taxonomy, Molecular Biology, Ecology, Pathology, and Control, Kluwer Academic Publishers, Dordrecht, The Netherlands, 578 pp.Zadoks J. C., Chang T. T., Konzak C. F., 1974 - A Decimal Code for the Growth Stages of Cereals. Weed Research, 14, 415 - 421.Wiese M.V., 1987 - Compendium of wheat diseases. American Phytopathological Society. pp. 124 pp.
Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
Patogenul care supraviețuiește în sămânța de grâu

Helmintosporioza lenticulară a grâului este o boală periculoasă care apare în culturile de grâu în anii când condițiile climatice sunt propice infecțiilor. În rândurile de mai jos sunt informații despre simptomatologia, biologia și strategia de combatere a acestei boli care poate produce pagube importante în producția de grâu.
Fungul Drechslera tritici – repentis a apărut în culturile de grâu din vestul României de prin anul 2002, dar din cauza tabloului simptomatic asemănător cu cel al fungului Septoria nodorum, a trecut neobservat (din cauza confuziei era raportat ca fiind Septoria nodorum). Identificarea corectă și prima raportare cu privire la acest patogen în Câmpia Banatului a fost făcută de către Cotuna & Popescu în anul 2007. În celelalte zone din țară patogenul era deja raportat.
Importanța economică și supraviețuirea patogenului
Helmintosporioza lenticulară este o boală gravă a grâului, capabilă să producă pagube cuprinse între 3% și 40% și chiar mai mult în anii extrem de favorabili. Infecțiile din timpul înfloritului sunt periculoase deoarece patogenul trece în sămânță. Cele mai mari pagube în producție se înregistrează atunci când infecțiile au loc în fazele de burduf și de înflorit [Shabeer & Bockus, 1988].
O sursă de inocul importantă este miceliul din seminţele infectate. Miceliul se dezvoltă în condiţii de temperatură cuprinse între 5 – 350C (la fel ca şi conidiile). Boabele sunt infectate sau predispuse la infecţie mai ales atunci când frunza stindard este afectată de boală şi când umiditatea este mare în perioada înspicatului. De altfel, ele pot fi infectate oricând de-a lungul perioadei de vegetaţie, însă predispoziţia maximă de îmbolnăvire este în timpul înspicatului. La suprafaţa cariopselor infectate apare simptomul de „red smudge” sau pată roşie – McMullen & Adhikari, 2009.
Fungul trăiește în mod saprofit pe resturile vegetale ale plantei gazdă, care constituie sursa principală de inocul primar. Pe paiele care rămân pe sol peste anotimpul de iarnă se formează pseudoteciile (forma telomorfă sau sexuată) cu asce şi ascospori.

Recunoașterea simptomelor
Simptomele macroscopice ale bolii apar de obicei în patru etape:
-
Stadiul de pată, când pe locul de infecţie iniţial după două zile apare o coloraţie închisă la culoare;
-
Pată înconjurată de cloroză, care apare la 4 – 6 zile după infecţie;
-
Necroza apare la 6 – 8 zile de la infecţie, iar clorozele se transformă în necroze şi în paralel începe sporularea;
-
Colapsarea frunzelor, acestea fiind afectate în întregime, iar sporularea este maximă.
Simptomele macroscopice sau externe apar prima dată pe frunzele bazale, sub forma unor puncte sau pete mici, circulare, de culoare maro deschis. Aceste pete sunt rezultatul infecţiilor primare, care au loc de regulă în lunile aprilie – mai. Cel mai mare pericol de infecţie este la grâul care s-a cultivat după grâu, datorită resturilor vegetale care rămân la suprafaţa solului. Frunzele bătrâne sunt primele infectate sau predispuse la infecţie.
Pe măsură ce patogenul evoluează, petele capătă formă ovală sau lenticulară, au culoare maro deschis şi sunt înconjurate de un halou gălbui (rezultatul infecţiilor secundare) – Schilder & Bergstrom, 1993; Cotuna & Popescu, 2008. În condiții favorabile de climă (nopţi umede şi calde), petele cresc în dimensiuni, se unesc iar pe frunze apar pete mari, cu aspect neregulat, la care se mai observă punctul iniţial de infecţie (aspect de necroză). Frunzele puternic atacate se usucă începând de la vârf. În faze avansate de infecţie, haloul sau zona clorotică din jurul petelor nu se mai observă (Wolf, 1991).
Pentru evitarea punerii unui diagnostic greșit (risc de confuzie cu Septoria nodorum), petele trebuie atent analizate pentru a vedea dacă pe ele apar suporţii negri ai sporilor. La mostrele colectate din câmp, conidiile nu se observă imediat, de aceea este necesară incubarea pe hîrtie umedă [Maraite et al., 1998; Cotuna, 2007].
A nu se confunda cu Septoria nodorum

Realizarea infecțiilor
Cercetările efectuate în ultimii ani în ceea ce priveşte epidemiologia acestei boli arată că, efectele cumulate ale temperaturilor ridicate, umidităţii relative mari şi a duratei de umectare a frunzelor mai mare de 12 ore (precipitaţii, rouă) conduc la apariţia şi dezvoltarea patogenului Drechslera tritici – repentis (Duveiller, 2004). Dezvoltarea patogenului este optimă la temperaturi cuprinse între 20 – 280C. Boala este asociată de regulă cu câmpurile pe care o cantitate mare de paie rămâne la suprafaţa solului (Bockus & Claassen, 1992).
Primele infecții sau infecțiile primare sunt realizate de ascosporii din pseudotecii care devin maturi primăvara. În condiții de umiditate, pe timp de noapte aceștia sunt eliberaţi din pseudotecii. Emisia de ascospori este maximă în lunile martie – aprilie şi are loc în condiţii de umiditate (şi roua este suficientă). Ascosporii sunt eliberaţi din asce la distanţe mici, de câţiva centimetri, spre deosebire de conidii care pot fi propagate pe distanţe mai mari cu ajutorul vântului (Rees & Platz, 1992; Murray et al., 2009).
Conidiile produc infecţiile secundare care afectează frunzele din etajele superioare (astfel apar şi pagubele în recoltă). Conidioforii se formează pe timp ploios, iar conidiile în timpul nopţilor calde şi umede la suprafaţa necrozelor de pe frunze şi sunt diseminate de vânt către plantele sau frunzele sănătoase. Dispersarea conidiilor are loc în timpul după – amiezei. Conidiogeneza, după cum am arătat mai sus, are loc în timpul nopţii, când umiditatea este mai ridicată şi alte suprafeţe purtătoare de conidii sunt uscate (Morrall & Howard, 1975; Platt & Morrall, 1980a, 1980b; Francl & Jordahl, 1992; Francl, 1997).
Sporii anamorfi pot fi dispersați pe kilometri întregi şi chiar zeci de kilometri (diseminare anemochoră), în zilele calde şi secetoase la temperaturi de 21 – 230C (optim). Ei germinează şi infectează grâul în condiţii variate de temperatură (5 – 350C), când frunzele sunt umede o perioadă de timp specifică, de aici rezidă și agresivitatea fungului.
În centrul petelor, punctul de infecție rămâne vizibil și după uscarea frunzelor

Managementul integrat al „helmintosporiozei lenticulare a grâului”
În cadrul sistemului de combatere integrată există o serie de măsuri care trebuie utilizate echilibrat pentru a ține sub control această boală.
Factorii care susțin infecțiile sunt:
-
Rotațiile scurte;
-
Lucrările minimale ale solului;
-
Densitatea mare a plantelor;
-
Rezistența la fungicide;
-
Soiurile sensibile;
-
Temperaturile ridicate;
-
Umiditatea relativă mare [Bockus & Claassen, 1992; Duveiller, 2004].
Măsuri profilactice
În general, helmintosporioza grâului este gravă în sistemele de cultivare în care lucrările solului sunt minimale sau nu se fac deloc (no tillage, minimum tillage) şi resturile vegetale rămân la suprafaţa solului. În sistemele convenţionale, boala este mai redusă din cauza îngropării resturilor vegetale. Se poate spune că factorii care influenţează intensitatea atacului sunt: lucrările minimale ale solului, irigarea, semănatul în afara epocii optime, fertilitatea scăzută a solului, monocultura (Sharma & Duveiller, 2003).
Prevenția constă în respectarea următoarelor măsuri agrofitotehnice:
-
Îngroparea resturilor vegetale sau îndepărtarea acestora pentru reducerea sursei de inocul (Diehl et al., 1982);
-
Utilizarea la semănat a soiurilor rezistente la boală este esențială;
-
Seminţele să fie libere de patogen;
-
Tratarea seminţelor înainte de semănat;
-
Respectarea asolamentului (asolament cu alte culturi care nu sunt gazde pentru patogen);
-
Rotaţia culturilor de cel puţin 3 – 4 ani;
-
Evitarea monoculturii;
-
Evitarea irigatului în perioada împăierii – înspicării (Docea & Severin, 1990);
-
Respectarea epocii optime de semănat.
Măsuri chimice
În cadrul strategiei de combatere, măsurile chimice dețin ponderea în prezent. Tratarea semințelor și aplicările foliare în timpul sezonului de vegetație sunt importante în combaterea helmintosporiozei lenticulare.
Combaterea chimică este dificilă atunci când infecţia s-a realizat deja, deoarece evoluţia acestei boli cu greu mai poate fi oprită de fungicide.
Simptomul de red smudge apare mai rar la Triticum aestivum și mai des la Triticum durum.Cel mai întâlnit simptom este de albire însoțit uneori de șiștăvire
Înainte de semănat se recomandă tratarea seminţelor. În Aplicația Pesticide 2.24.8.2 (2024) este doar un singur fungicid omologat la triticale, dar nu și la grâu, pe bază de difenoconazol + fludioxonil. Cu siguranță, fungicidele omologate pentru alți patogeni din sămânță controlează și patogenul Pyrenophora tritici repentis. În trecut erau mai multe produse de protecția plantelor omologate. Se pare că au fost scoase, deși apar omologate la alte helmintosporioze (ale orzului, de exemplu).
Fungul Drechslera tritici – repentis este greu combătut cu fungicide după ce frunzele sunt infectate. Este indicată aplicarea fungicidelor la apariţia primelor sporulări (stadiul EC 32 – 37). Deoarece sporularea are loc la suprafaţa necrozelor, analizele pentru stabilirea diagnosticului se vor face la frunzele bazale care de obicei sunt şi cele mai afectate (Kremer & Hoffman, 1993; Habermeyer & Gerhard, 1997).
Se recomandă ca tratamentele chimice să se efectueze în urma controlului fitosanitar al culturilor de grâu. Dacă se constată că sunt simptome, trebuie intervenit rapid pentru stoparea infecției, deoarece patogenul este destul de greu de ținut sub control din momentul în care infecția s-a realizat. În anii deosebit de favorabili, ciuperca și-a făcut apariția în culturi chiar din a treia decadă a lunii martie (în Banat).
Fungicidele ce pot fi utilizate în combatere sunt: Protioconazol + tebuconazol; Fenpropidin; Tebuconazol; Protioconazol; Bixafen + tebuconazol; Azoxistrobin; Metconazol; Azoxistrobin + protioconazol; Benzovindiflupir; Tebuconazol + trifloxistrobin; Protioconazol + spiroxamină + trifloxistrobin; Fluxapyroxad + piraclostrobin; Proquinazid + protioconazol; Bixafen + protioconazol; Bixafen + spiroxamină + trifloxistrobin; Tetraconazol; Fluxapyroxad + metconazol; Piraclostrobin;
Mefentrifluconazol + metrafenonă + piraclostrobin; Fluxapyroxad + metconazol [APLICAȚIA PESTICIDE 2.24.8.2, 2024].
În prezent, se recomandă utilizarea cu prudenţă a pesticidelor, aplicarea tratamentelor la momentul optim sau când se întrunește pragul economic de dăunare, respectarea dozelor recomandate de producători. Procedând așa, numărul de tratamente se va reduce simțitor, la fel și costurile.
Wolf & Hoffman (1994), în urma cercetărilor efectuate în Bavaria au stabilit două praguri pentru aplicarea unui tratament:
-
Când conidiile sunt găsite pe 5% din frunze;
-
Când simptomele apar la peste 5% din frunzele din etajul superior al plantelor.
Măsuri biologice
Tratarea semințelor cu agentul biologic Bacillus subtillis a dat rezultate foarte bune în controlul acestui patogen. Fungii Limonomyces roseipellis și Laetisaria arvalis (basidiomicotine) aplicați pe paiele de grâu în prezența umidității au redus sursa de inocul în procente ridicate. În urma aplicării s-a constatat că fungul Limonomyces roseipellis a redus inoculul cu 50 până la 80 și chiar 99% în anumite situații [Pfender et al., 1993].
BibliografieBochus W. W., Classen M. M., 1992 – Effects of crop rotation and residues management practices on severity of tan spot of winter wheat. Plant Dis., 76: 633 – 636.Cotuna Otilia, Popescu Gheorghe, 2008 – Helmintosporioza lenticulară, o boală nouă la grâul cultivat în Câmpia Banatului, Rev. Sănătatea plantelor, 2008;Diehl J. A., Tinline R. D., Kochhann R. A., Shipton P. J., Rovira A. A., 1982 – The effect of fallow periods on common root rot of wheat in Rio Grande do Sul, Brazil. Phytopathology, 72: 1297 – 1301.Docea E., Severin V., 1990 – Ghid pentru recunoaşterea şi combaterea bolilor plantelor agricole, vol. I, Ed. Ceres, Bucureşti, 320 p.Duveiller E., 2004 – Controlling foliar blight of wheat in the rice – wheat systems of Asia. Plant Dis. 88: 552 – 556.Francl L. J., Jordahl Y. G.,1992 – Spike inoculation of durum wheat with Pyrenophora tritici – repentis and seed symptomology. Proceeding of the second International Tan Spot Worshop, Fargo, ND.Francl L., 1997 – Local and mesodistance dispersal of Pyrenophora tritici – repentHabermeyer J., Gerharg M., 1997 – Pilzkrankheiten und Schasymptome im Getreidebau, BASF, p. 41 – 45.Kremer M., Hoffman G. M., 1993 – Effekte von Blattinfektionen durch Drechslera tritici – repentis auf den kohlenhydrat und stickstoffhaus halt von weizenpflanzen.Maraite H., Di Zinno T., Longrée H., Daumerie V., Duveiller E., 1998 – Fungi associated with foliar blight of wheat in warm areas. In E. Duveiller, H. J. Dubin, J. Reeves & A. McNab, eds. Proc. Int. Workshop Helminthosporium Diseases of wheat: Spot Blotch and tan spot, CIMMYT, El Batan, Mexico, 9 – 14 Feb. 1997, p. 293 – 300.McMullen Marcia, Tika Adhikari, 2009 – Fungal leaf spot Diseases of wheat: Tan spot, Stagonospora nodorum blotch and Septoria tritici blotch. PP – 1249 (revised), disponibil pe www.ag.ndsu.edu.Morrall R. A. A., Howard R. J., 1975 – The epidemiology of leaf spot disease in a native prairie 11 Airborne spore populations of Pyrenophora tritici – repentis. Can J. Bot., 53: 2345 – 2353.Murray T. D., Parry D. W., Cattlin N. D., 2009 – Diseases of small grain cereal crops, Manson Publising Ltd, London, U. K., 142 pp.Pflender W. F., Zhang W., Nus A., 1993. Biological control to reduce inoculum of the tan spot pathogen Pyrenophora tritici-repentis in surface - borne residues of wheat fields. Phytopathology, 83: 371 - 375.Platt H. W., Morrall R. A. A., 1980a – Effects of light intensity and relative humidity on conidiation of Pyrenophora tritici – repentis. Can J. Plant Pathol., 2: 53 – 57.Platt H. W., Morrall R. A. A., 1980b – Effects of wind speed and humidity on conidiation of Pyrenophora tritici – repentis. Can J. Plant Pathol., 2: 58 – 64.Rees R. G., Platz G. J., 1992 – Tan spot and its control – some Australian experiences. In L. J. Francl, J. M. Krupinsky, M. P. Mc.Mullen, eds. Advances in tan spot research, p. 1 – 15, NDSU Agric. Exp. Sta. Publ. 146 pp.Shabeer A., Bockus W. W., 1988. Tan spot effects on yield and yield components relative to growth stage in winter wheat. Plant Disease, 72: 599 - 602.Sharma R. C., Duveiller E., 2003 – Effect of stress on Helminthosporium leaf blight in wheat. Pages 140 – 144 in: Proc. 4th Int. Wheat Tan spot and spot blotch Workshop. J. B. Rasmussen, T. L. Friesen and S. Ali, eds. North Dakota State University, Fargo;Schilder A. M. C., Bergstrom G., 1993 – Tan spot. In S. B. Mathur & B. M. Cunfer, eds. Seedborne diseases and seed health testing of wheat, p. 113 – 122. Copenhagen, Denmark, Jordburgsforlaget;Wolf P., 1991 – Biology, epidemiology, Schadrelevantz, konzeption für eine integrierte Bekämpfung von Drechslera tritici – repentis (Died.) Shoem., (Perfektstadium Pyrenophora tritici – repentis (Died.) Drechs., dem Erreger einer Blattfleckenkrankheit. Dissertation TU München;Wolf P. F. J., Hoffmann G. M., 1994. Decision model based on the epidemiology for integrated control of Drechslera tritici-repentis (Died.) Shoem., (teleomorph Pyrenophora tritici-repentis (Died.) Drechs.). Gesunde Pflanzen 46, 164–173.***APLICAȚIA PESTICIDE 2.24.8.2, 2024.
Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
Boala care a distrus și în 2024 multe culturi de floarea-soarelui, putrezirea cărbunoasă a rădăcinilor și tulpinilor

Readuc în atenția fermierilor Macrophomina phaseolina, patogenul care și în acest an a produs pagube în culturile de floarea-soarelui. Seceta a fost favorabilă dezvoltării acestui fung (iubește căldura).
În 2024, seceta, arșița și fungul Macrophomina phaseolina au distrus multe culturi de floarea-soarelui, mai ales în sudul României. De asemenea, și în Banat fungul este prezent în culturile de floarea-soarelui. În această perioadă putem observa plantele complet uscate și căzute la sol. Există zone în țară unde plantele de floarea-soarelui sunt încă verzi și aparent sănătoase. În Timiș am observat culturi care sunt înflorite și au toate frunzele verzi. Este vorba de acele culturi care au fost semănate la sfârșitul lunii mai și au scăpat de arșiță. Macrophomina phaseolina este prezentă și în aceste sole (observații realizate la data de 6 august 2024). De ce acest fung câștigă teren tot mai mult în România? La răspuns trebuie să ne gândim cu toții. Care sunt cauzele? Le veți afla parcurgând acest material.

Care sunt factorii care au condus la creșterea sursei de inocul a fungului Macrophomina phaseolina în solurile din România? Am identificat acest patogen în Banat prin anul 2021 la cererea unui fermier care nu știa cu ce se confruntă. În acel an, fungul a pus la pământ mai multe hectare de floarea-soarelui în ferma de unde proveneau probele. De atunci și până în prezent am observat că fungul continuă să producă pagube în fiecare vară, iar incidența și intensitatea atacului sunt tot mai ridicate.

Factorii de risc
Factorii care contribuie la extinderea acestui patogen în România sunt:
-
Lipsa apei din sol - predispune plantele de floarea-soarelui la atacul agenților patogeni sistemici care distrug și blochează vasele [Vear, 2016; Debaeke et al., 2017].
-
Creșterea temperaturilor peste 280C. Vremea caldă și secetoasă stimulează patogenul Macrophomina phaseolina. După Sarova et al. (2003), condițiile de vreme caldă și uscată (temperaturi cuprinse între 28 - 300C și lipsa apei din sol) favorizează instalarea fungului. Temperatura, umiditatea atmosferică și cea disponibilă sunt foarte importante în realizarea infecțiilor cu Macrophomina phaseolina. După Marquez et al. (2021), microscleroții germinează la temperaturi cuprinse între 30 - 350C.
-
Caracterul invaziv al patogenului phaseolina (reiese din cele mai multe studii analizate). Pe lângă asta, numărul mare de plante gazdă, distribuția la nivel global, schimbările climatice arată că fungul prezintă importanță deosebită pentru viitorul culturii de floarea-soarelui și nu numai [Cotuna et al., 2022].
-
Condițiile de sol, înrădăcinarea defectuoasă a plantelor, carențele de bor (patogenul se instalează cu ușurință pe plantele afectate de fiziopatii) - Popescu, 2005. Același autor arată că fungul infectează în general plantele cu afecțiuni fiziopatice, la care creșterea rădăcinii principale este stopată, iar rădăcinile secundare încep să îmbătrânească. La aceste plante, sistemul radicular va fi ocupat de Fusarium sp., dar și de alte ciuperci care pregătesc astfel țesuturile radiculare pentru infecția cu Macrophomina phaseolina. Aproape întotdeauna, pe rădăcinile atacate de fung se observă micelii albe - rozii specifice fungului Fusarium sp.
-
Densitatea ridicată, rănile mecanice, atacul insectelor favorizează instalarea patogenului [Shiekh & Ghaffar, 1984; Ahmed et al., 1991].
-
Prezența microscleroților în sol în cantitate mare. Macrophomina phaseolina rezistă în sol sub formă de microscleroți, pe resturile vegetale, dar și în masa de semințe [EPPO, 2000; Csüllög et al., 2020; Popescu, 2005; Docea & Severin, 1990]. Microscleroții pot supraviețui în sol de la doi până la 15 ani [Baird et al., 2003; Gupta et al., 2012; Csüllög et al., 2020].
-
Lucrările minimale ale solului.
-
Rotațiile scurte.
Analizând factorii de risc, cu ușurință ne dăm seama că, creșterea temperaturilor în zonele cu climat temperat, însoțită de lipsa precipitațiilor ar putea crea probleme deosebite în culturile de floarea-soarelui în viitor, făcându-le vulnerabile la atacul patogenului M. phaseolina [Debaeke et al., 2017].

Cum putem lupta cu acest patogen?
Deoarece prin metode chimice nu putem controla patogenul, pierderile pot fi evitate dacă ținem cont de câteva măsuri profilactice:
-
Cultivarea hibrizilor rezistenți sau toleranți la boală.
-
Irigarea culturilor în condiții de secetă și temperaturi ridicate.
-
Distrugerea resturilor vegetale infectate (sunt pline de microscleroți).
-
Înființarea culturilor în soluri cu textură corespunzătoare.
-
Respectarea rotației culturilor. Cu privire la rotație, nu întotdeauna rezultatele sunt cele scontate din cauza polifagiei ciupercii, care are capacitatea de a infecta peste 300 de plante cultivate și buruieni [Francl et al., 1988; Hafeez & Ahmad, 1997; EPPO, 2000; Popescu, 2005].
-
Utilizarea la semănat de sămânță liberă de microscleroți, lucrări ale solului de calitate superioară, igiena culturală [Docea & Severin, 1990].

Mai multe informații cu privire la recunoașterea simptomelor, biologia și managementul integrat al fungului Macrophomina phaseolina găsiți în articolele pe care le-am documentat, scris și publicat de-a lungul timpului:
-
https://revistafermierului.ro/din-revista/protectia-plantelor/item/5111-putrezirea-carbunoasa-a-radacinilor-si-tulpinilor-de-floarea-soarelui.html.

Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna
Abonamente Revista Fermierului – ediția print, AICI!
Fuzarioza ar putea produce probleme în unele zone, la culturile de cereale păioase

Aduc în atenția fermierilor fungul Fusarium graminearum care produce boala numită „albirea și înroșirea spicelor”. În zonele din țară unde au căzut precipitații în perioada înfloritului, iar temperaturile au fost favorabile realizării infecțiilor, este posibil să apară fuzarioza.
Schimbările climatice din ultimii ani au influențat pozitiv dezvoltarea fungilor din genul Fusarium, favorizând apariția epidemiilor la grâu. Cea mai recentă epidemie de Fusarium la cerealele din Câmpia Banatului (și nu numai) a fost în anul 2019, an în care calitatea a fost foarte scăzută, în principal din cauza prezenței micotoxinelor fusariene în cantități care au depășit limitele permise.
Pierderile produse de F. graminearum la grâu în anii epidemici pot fi uriașe. Pagubele se datorează în mare parte sterilității spicelor, MMB-ului scăzut (masa a o mie de boabe), dar mai ales prezenței micotoxinelor în cariopse.
Fusarium graminearum este un patogen deosebit de periculos al cerealelor deoarece produce micotoxine încadrate în două clase chimice: trichothecene și zearalenon. Dintre trichothecene amintim: vomitoxina (deoxynivalenol sau DON), micotoxina T – 2, diacetoxyscirpenol (DAS), monoacetoxyscirpenol (MAS) şi nivalenol. Aceste micotoxine sunt iritanţi puternici şi au fost asociate atunci când sunt consumate cu simptome ca: vomă, refuzul hranei şi posibil ulcer gastric. Cele mai semnificative trichothecene sunt toxina T – 2 şi deoxynivalenolul, care apar în cantităţi destul de mari la cereale. Zearalenonul face parte din a doua clasă chimică de toxine produse de F. graminearum. Când este consumat de animale este asociat cu probleme de reproducere, cum sunt: avorturile, căldurile false, reabsorbţia fetusului şi a mumiilor [Cotuna & Popescu, 2009].
Fusarium și Alternaria sp. (foto din anul 2023)

În Câmpia Banatului, în anul 2023 au existat lanuri infectate, însă incidența spicelor atacate a fost mai scăzută, la fel și intensitatea. De la epidemia de Fusarium graminearum din anul 2019, putem aprecia că acest patogen nu a mai creat probleme deosebite în Banat, deoarece nu s-au întrunit condițiile climatice (precipitații continue și temperaturi moderate). Vom vedea ce va aduce această primăvară.
Prin intermediul acestui articol venim în sprijinul dumneavoastră cu informații despre tabloul simptomatic al bolii, biologia, epidemiologia și „combaterea” patogenului Fusarium graminearum. Aceste informații vă vor ajuta în viitor să vă protejați din timp culturile.

Micotoxinele fusariene, pericol pentru sănătatea oamenilor și animalelor
În fuzarioza grâului pot fi implicate mai multe specii de Fusarium. Studii numeroase arată că fuzarioza spicelor de grâu poate fi produsă de Fusarium graminearum, Fusarium culmorum, Fusarium nivale, Fusarium poae, Fusarium sporotrichioides [Miller, 1994; Lidell, 2003; Wegulo, 2012; Zrcková et al., 2019]. Dintre speciile menționate, Fusarium graminearum este prezentă în regiunile temperate cu climat mai cald, comparativ cu Fusarium culmorum care preferă zonele mai reci [Wang & Miller, 1988; Snijders & Perkowski, 1990; Miller et al., 1991; Miller, 2002]. În Câmpia Banatului, specia predominantă care produce infecții la spic este F. graminearum [Cotuna et al., 2013; Cotuna et al., 2022].
Dintre speciile de Fusarium producătoare de DON, F. graminearum este considerată cea mai importantă [Paraschivu et al., 2014; Paul et al., 2005; Anon, 1993c]. Deoxynivalenolul (DON) aparține familiei chimice de sequiterpene, fiind derivat din trichodiene (precursorul biochimic al tuturor trichothecenelor). DON - ul este foarte stabil din punct de vedere chimic. Semințele infectate de Fusarium conțin întotdeauna și micotoxine fusariene. Dintre acestea, DON - ul a fost găsit frecvent în cantități mari [McMullen et al., 1997]. După Wegulo (2012), cu cât procentul de boabe fusariate este mai mare cu atât și cantitatea de DON va fi mai ridicată. De altfel, marea majoritate a cercetătorilor corelează prezența deoxynivalenolului în cariopse cu intensitatea atacului din câmp și procentul de boabe fusariate [Cowger & Arellano, 2013]. Dacă ajunge în hrana oamenilor, deoxynivalenolul poate produce intoxicații alimentare, care se manifestă prin greață, vărsături, diaree, dureri de cap, dureri abdominale, febră etc [Lidell, 2003; Sobrova et al., 2010].
Până în acest an, limita maximă de DON admisă de legislația europeană în cerealele neprocesate era de 1250 ppb (1,25 ppm) [Commission Regulation (EC) No 1881/2006].
În Regulamentul (UE) 2024/1022 al CE din 8 aprilie 2024, de modificare a Regulamentului (UE) 2023/915 cu privire la nivelurile maxime de deoxinivalenol în produsele alimentare, limita maximă de DON permisă la cerealele neprocesate a scăzut la 1000 ppb. Noile reglementări intră în vigoare începând cu data de iulie 2024 și nu se aplică retroactiv.

A doua micotoxină importantă produsă de Fusarium graminearum este „toxina T - 2”, care apare în cantități semnificative la cereale, alături de deoxynivalenol [Annon, 1993b]. Intoxicația se manifestă prin simptome de febră, vomă, convulsii, anemie, inflamații acute ale aparatului digestiv.
Alt metabolit toxic produs de fungul F. graminearum este zearalenona (ZON). Zearalenona apare la grâul fusariat alături de DON și T - 2. Această toxină afectează eficiența reproductivă, nu și pofta de mâncare. Sindromul estrogenic ce apare în urma ingerării de hrană contaminată se caracterizează prin: umflarea glandelor mamare, hipertrofia uterină, umflarea vulvei, infertilitate [Marasas, 1991]. Cei mai sensibili sunt porcii.
Limitele maxime admise de ZON și T - 2 în grâul neprocesat sunt de 100 ppb. Cele trei micotoxine, DON, ZON și T - 2 nu sunt considerate carcinogenice. Zearalenona nu se transmite prin lapte sau alte produse lactate.
Factorii de risc pentru apariția infecțiilor
Risc crescut de infecții cu Fusarium graminearum se înregistrează în anii când se întrunesc următorii factori:
-
Temperaturi optime pentru realizarea infecțiilor. După Anderson (1948), temperatura optimă pentru realizarea infecțiilor este de 250C, indiferent de cât timp durează umezeala. După De Wolf et al. (2003), contează durata în ore a temperaturilor cuprinse între 15 - 300C, înainte cu șapte zile de înflorit. În condiții de vreme caldă cu temperaturi cuprinse între 25 - 300C și umiditate continuă, simptomele de Fusarium la spic (albire) pot apărea în 2 - 4 zile de la realizarea infecției [Wegulo, 2012]. Astfel, o cultură aparent sănătoasă, brusc poate să prezinte simptome de boală;
-
Precipitațiile. Precipitațiile continue dinainte de înflorit și în timpul dezvoltării cariopselor favorizează acumularea de cantități mari de DON în cereale. Cantitățile de precipitații din lunile mai și iunie predispun cerealele la infecția cu Fusarium. Perioadele în care grâul poate fi infectat sunt la înflorit sau imediat după înflorit [Hernandez Nopsa et al., 2012; Wegulo, 2012]. De Wolf et al. (2003) arată importanța duratei în ore a precipitațiilor înainte cu șapte zile de înflorit;
-
Umiditatea relativă a aerului (UR%). Cu cât expunerea la umezeală este mai îndelungată, intensitatea atacului la spic crește. Chandelier et al. (2011), într-un studiu efectuat pe o perioadă de șapte ani, arată o corelație puternică între umiditatea relativă medie de peste 80% și cantitatea de DON acumulată în cariopse;
-
Tehnologiile practicate în prezent de către fermieri pot influența pozitiv infecțiile cu Fusarium, cât și acumularea de micotoxine. Sistemele de cultivare „minimum tillage” sau „no tillage” (utile pentru conservarea solului), densitățile mari practicate, lipsa rotației, au dus la creșterea sursei de inocul în resturile vegetale ce rămân la suprafața solului [Unger, 1994; Watkins, 1994; Matei et al., 2010];
-
Soiurile sensibile.
Recunoașterea simptomelor
Fusarium graminearum poate ataca plantele de cereale păioase pe tot parcursul perioadei de vegetație, dacă condițiile climatice preferate se întrunesc.
Tabloul simptomatic al bolii se prezintă după cum urmează:
-
Plăntuţele care provin din seminţe infectate se îngălbenesc şi în cele din urmă putrezesc;
-
În faza de înfrăţire, rădăcinile şi coletul sunt brunificate din cauza infecţiilor realizate de miceliul şi clamidosporii din sol. Plantele atacate continuă să vegeteze slab şi vor forma spice sterile;
-
Forma cea mai gravă de atac este după înspicare. Spicele, iniţial se albesc parţial (câteva spiculeţe) sau total, apoi se înroşesc şi se acoperă cu un înveliş micelian, alb – roz sau alb – rubiniu, uneori portocaliu - somon, pe care se observă sporodochiile ciupercii (forma imperfectă). Pe spicele înroşite (pe palee, ariste sau boabe) se observă puncte negre care sunt periteciile ciupercii (forma perfectă). Cariopsele infectate sau fuzariate rămân mici, zbârcite, cenuşii sau rozii iar germinaţia şi puterea de străbatere va fi slabă [Popescu, 2005].

Ciclul de viață
Fusarium graminearum este agentul etiologic dominant al fuzariozei spicului la cerealele păioase cultivate în România. Ciuperca rezistă în resturile de plante vegetale, în sol și în semințe. Vremea umedă prelungită în timpul perioadei de vegetație favorizează creșterea și sporularea ciupercii. Sporii ciupercii sunt purtați de vânt și de picăturile de apă pe spicele de grâu. Grâul este susceptibil a fi infectat în perioada înfloritului și când cariopsele încep să se formeze [Popescu, 2005].
Fusarium graminearum rezistă în sol sub formă de miceliu saprofit, clamidospori şi peritecii. O sursă importantă de transmitere este sămânţa infectată din care ies plăntuţe bolnave care mor (infecţie sistemică). Infecţiile primare pot fi realizate de micelii sau clamidosporii din sol dar şi de ascosporii şi conidiile care ajung pe părţile aeriene ale plantelor. După realizarea infecției, miceliul care se dezvoltă intracelular va intra în sporogeneză, formându-se astfel conidiile ce asigură infecţiile secundare (foarte păgubitoare mai ales în perioada înfloritului) – Popescu, 2005.
Dezvoltarea acestui patogen este favorizată de vremea umedă (umiditatea aerului peste 90%, prezenţa ploilor) şi de temperaturile moderate (peste 200C) şi apoi de factorii agrofitotehnici (monocultura, solurile acide, azotul în exces, semănatul des, sensibilitatea soiurilor).
Infecţia continuă şi în depozite. Contaminarea cu micotoxinele produse de F. graminearum este asociată cu amânarea excesivă a recoltatului şi cu depozitarea cerealelor umede. Acumularea de micotoxine este masivă la temperaturi de 21 – 290C şi la o umiditate a boabelor de peste 20%.
Managementul integrat al fuzariozei grâului
Putem combate sau nu fuzarioza la cereale? O întrebare la care este greu de răspuns. Măsurile din cadrul sistemului de combatere integrată pot ține sub control destul de puțin fuzarioza dar nu întotdeauna ne feresc de infecții. De ce? Pentru că orice măsuri am respecta, condițiile climatice sunt esențiale în realizarea infecțiilor.
Atac la cariopse. Stanga, cariopse fusariate, dreapta cariopsă aparent sănătoasă (foto din anul 2023)

Măsuri profilactice
Măsurile de profilaxie sunt foarte importante dar nu ne feresc de infecții dacă condițiile climatice sunt favorabile patogeniei. Totuși, respectarea lor ne poate ajuta, în sensul că vom avea o rezervă mai mică în sol de inocul. În acest sens, este bine ca fermierii să respecte următoarele măsuri:
-
Cultivarea de soiuri adaptate climei locale şi zonei unde vor fi cultivate.
-
Cultivarea unor soiuri care tolerează mai bine patogenul. Despre rezistență totală nu putem discuta. Rezistența soiurilor de grâu la infecția cu Fusarium este foarte importantă și intens studiată astăzi. Sunt descrise până acum cinci tipuri de rezistență: tipul I - rezistența la infecția inițială (reacții de apărare); tipul II - rezistența la răspândirea agentului patogen în țesutul infectat; tipul III - rezistența la infecție a semințelor; tipul IV - toleranța la infecție; tipul V - rezistența la micotoxine [Mesterhazy, 1995; Ma et al., 2009; Kosaka et al., 2015; Zhang et al., 2020]. După Bai & Shaner (2004), crearea unor soiuri cu rezistență la Fusarium poate fi o strategie foarte bună pentru controlul acestei boli. În SUA, preocupări de ameliorare a grâului pentru rezistența la Fusarium sp. există de prin anul 1929. Un studiu din 1963 arată că, după un ciclu de cercetari de nouă ani, toate plantele de grâu pot fi infectate în proporție mai mare sau mai mică [Schroeder & Christensen, 1963].
-
Controlul dăunătorilor în lanurile de cereale nu trebuie neglijat, deoarece se ştie că favorizează infecţiile cu Fusarium graminearum.
-
Densităţile mari trebuie evitate.
-
Fertilizarea cu azot şi alte substanţe nutritive să se facă în mod echilibrat.
-
Rotaţia culturilor trebuie respectată, deoarece s-a constatat că reduce riscul de contaminare cu micotoxine produse de ciuperca Fusarium graminearum.
-
Resturile vegetale să fie îngropate prin intermediul arăturii.
-
Recoltarea la timp, uscarea la 24 de ore de la recoltare şi supravegherea umidităţii boabelor la depozitare [Cotuna & Popescu, 2009].
Dacă aceste măsuri sunt respectate, sursa de inocul va fi diminuată, NU şi eliminată.

Măsuri chimice
În funcție de condițiile climatice, tratamentele chimice pot fi eficiente sau nu. Tratarea semințelor înainte de semănat este esențială în prevenirea primelor infecții.
În România sunt omologate următoarele substanțe pentru tratarea semințelor de cereale păioase: Triticonazol; Tebuconazol; Fludioxonil + teflutrin (insecticid); Fludioxonil + protioconazol + tebuconazol; Fludioxonil; Difenoconazol + fludioxonil; Difenoconazol + fludioxonil + tebuconazol; Difenoconazol; Fludioxonil + fluxapyroxad + triticonazol; Ipconazol; Fluxapyroxad; Fludioxonil + sedaxan; Difenoconazol + fludioxonil + sedaxan; Bixafen + tebuconazol [după Aplicația PESTICIDE 2.24.3.1, 2024].
Tratamentele din vegetație
La modul general, în literatura de specialitate se recomandă două tratamente în timpul sezonului de vegetație, după cum urmează: primul tratament la începutul înspicării; iar al doilea tratament la sfârșitul înfloritului.
Studiile efectuate pentru stabilirea momentelor optime de efectuare a tratamentelor (când au eficacitate maximă) recomandă următoarea strategie:
-
Tratament la BBCH 59 - când grâul nu este înflorit - eficiență ridicată.
-
Tratament la BBCH 63 - 65 - început înflorit, moment optim pentru bolile spicului în general.
-
Tratament la BBCH 69 - sfârșit înflorit - nu se recomandă (prea târziu pentru tratament) - se poate aplica doar în situații grave cu risc de infecții secundare când sunt ploi continue după înflorit.
Pentru tratamentele în vegetație sunt omologate următoarele substanțe: Azoxistrobin; Tebuconazol; Metconazol; Azoxistrobin + protioconazol; Protioconazol + tebuconazol; Azoxistrobin + tebuconazol; Kresoxim - metil + mefentrifluconazol; Benzovindiflupir + protioconazol; Benzovindiflupir; Protioconazol; Protioconazol + spiroxamină + tebuconazol; Ciprodinil; Fenpropidin; Difenoconazol + tebuconazol; Tebuconazol + trifloxistrobin; Protioconazol + spiroxamină + trifloxistrobin; Protioconazol + trifloxistrobin; Boscalid + protioconazol; Fluxapyroxad + piraclostrobin; Mefentrifluconazol + piraclostrobin; Bromuconazol + tebuconazol; Proquinazid + protioconazol [după Aplicația PESTICIDE 2.24.3.1, 2024].
Fungicidele omologate trebuie utilizate doar în dozele recomandate de producători. Nu măriți dozele. Mărirea dozelor duce la apariția fenomenului de rezistență, iar rezistența la pesticide este o problemă mare a agriculturii moderne.
Tratamentele trebuie efectuate doar în zilele în care nu bate vântul și temperaturile nu sunt ridicate. Dacă după efectuarea tratamentelor intervin ploi, va trebui să repetați. Este foarte important să fie respectați timpii de pauză până la recoltat. Fungicidele utilizate la cereale au timpi de pauză destul de mari, începând de la 35 până la 50 zile.
Măsuri biologice
Combaterea biologică este foarte rar utilizată în combaterea fuzariozei la grâu și nu numai. De interes sunt antibioticele produse de bacterii (Bacillus subtilis) și fungi (Penicillium, Trichoderma, Trichothecium): fitobacteriomicina, nifimicina, fitoflavina, lavendromicina, trichotecina [Popescu, 2005].
În prezent, există un produs biologic omologat în România pe bază de Pythium oligandrum (M1 x 106 oospores/g Pythium oligandrum) pentru tratarea fuzariozei în perioada de vegetație. Tratamentele cu agenți biologici trebuie efectuate preventiv, nu curativ.
De reținut, recoltele contaminate cu micotoxine fusariene nu pot fi destinate nici pentru panificaţie, nici pentru hrana animalelor, din cauza intoxicaţiilor grave pe care le produc.
Bibliografie
Andersen, A. L., 1948. The development of Gibberella zeae head blight of wheat. Phytopathology, 38, 599 – 611.Anon, 1993b. In IARC Monographs on the evaluation of carcinogenic risk to humans, vol. 56, International Agency for Research an Cancer, Lyon, France, pp. 467 - 488.Anon, 1993c. In IARC Monographs on the evaluation of carcinogenic risk to humans, vol. 56, International Agency for Research an Cancer, Lyon, France, pp. 397 - 444.Bai, G., Shaner, G., 2004. Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 42: 135 - 161.Chandelier, A., Nimal, C., André, F., Planchon, V., Oger, R., 2011. Fusarium species and DON contamination associated with head blight in winter wheat over a 7-year period 92003–2009) in Belgium. Eur. J. Plant Pathol., 130, 403 – 414.Cotuna, O., Sărățeanu, V., Durău, C., Paraschivu, M., Rusalin, G., 2013. Resistance reaction of some winter wheat genotipes to the attack of Fusarium graminearum L. Schw. in the climatic conditions of Banat plain, Research Journal of Agricultural Science, 45 (1), p. 117 - 122.Cotuna O., Paraschivu M., Sărăţeanu V., Partal E., Durău C. C., 2022. Impact of Fusarium head blight epidemics on the mycotoxins’ accumulation in winter wheat grains, Emirates Journal of Food and Agriculture, 34 (11), 949 - 962.Cotuna O., Popescu G., 2009. Securitatea și calitatea produselor vegetale, siguranța vieții, Editura Mirton, Timișoara, 327 p..Cowger, C., Arellano, C., 2013. Fusarium graminearum infection and deoxynivalenol concentrations during development of wheat spikes. Phytopathology 103: 460 - 471.De Wolf, E. D., Madden, L. V., Lipps, P. E., 2003. Risk assessment models for wheat Fusarium head blight epidemics based on within-season weather data. Phytopathology, 93, 428 – 435.Hernandez Nopsa, J., Baenziger, P. S., Eskridge, K. M., Peiris, K. H. S., Dowell, F. E., Harris, S. D., Wegulo, S. N., 2012. Differential accumulation of deoxynivalenol in two winter wheat cultivars varying in FHB phenotype response under field conditions. Can. J. Plant Pathol. 34, 380 – 389.Kosaka, A., Manickavelu, A., Kajihara, D., Nakagawa, H., Ban, T., 2015. Altered gene expression profiles of wheat genotypes against Fusarium head blight. Toxins 72: 604 - 620.Liddell, C. M., 2003. Systematics of Fusarium species and allies associated with Fusarium head blight. In Fusarium Head Blight of Wheat and Barley; Leonard, K. J., Bushnell, W. R., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2003; pp. 35 – 43.Ma, H., Ge, H., Zhang, X., Lu, W., Yu, D., Chen, H., Chen, J., 2009. Resistance to Fusarium head blight and deoxynivalenol accumulation in Chinese barley. J. Phytopathology, 157, 166 – 171.Marasas, W. F. O., 1991. In Mycotoxins and Animal Foods (J. E., Smith, and R. S., Henderson, editors), CRC Press, Inc., pp. 119 - 139.Matei, G., Păunescu, G., Imbrea, F., Roşculete E., Roşculete, C., 2010. Rotation and fertilization - factors in increasing wheat production and improving the agro productive features of the brown reddish soil from central area of Oltenia, Research Jurnal Of Agricultural Science, Vol. 42 (1). USAMVB Timișoara, pag. 182 - 189.Mesterhazy, A. I., 1995. Types and components of resistance to Fusarium head blight of wheat. Plant breeding 114 5: 377 - 386.McMullen, M., Jones, R., Gallenberg, D., 1997. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 81:1340 - 1348.Miller, J. D., Greenhalgh, R., Wang, Y., Lu, M., 1991. Trichothecene chemotypes of three Fusarium species. Mycologia, 83, 121 – 130.Miller, J. D., 1994. Epidemiology of Fusarium ear diseases of cereals. In Mycotoxins in Grain. Compounds Other than Aflatoxin; Miller, J. D., Trenholm, H. L., Eds.; Eagan Press: St. Paul, MN, USA, 1994; pp. 19 – 36.Miller, J. D., 2002. Aspects of the ecology of Fusarium toxins in cereals. In Mycotoxins and Food Safety; DeVries, J. W., Trucksess, M. W., Jackson, L. S, Eds.; Kluwer Academic/Plenum Publishers: New York, USA, pp. 19 – 28.Paraschivu, M., Cotuna O., Paraschivu M., 2014. Integrated disease management of Fusarium head blight, a sustainable option for wheat growers worldwide, Annals of the University of Craiova - Agriculture, Montanology, Cadastre Series, vol. XLIV, p. 183 - 187.Paul, P. A., Lipps, P. E., Madden, L. V., 2005. Relationship between visual estimates of Fusarium head blight intensity and deoxynivalenol accumulation in harvested wheat grain: a meta-analysis. Phytopathology 95:1225 - 1236.Popescu G., 2005. Tratat de patologia plantelor, vol. II Agricultură, Editura Eurobit, 341 p..Snijders, C. H. A., Perkowski, J., 1990. Effects of head blight caused by Fusarium culmorum on toxin content and weight of wheat kernels. Phytopathology, 80, 566 – 570.Sobrova, P., Adam, V., Vasatkova, A., Beklova, M., Zeman, L., Kizek, R., 2010. Deoxynivalenol and its toxicity. Interdisc. Toxicol., 3, 94 – 99.Schroeder, H. W., Christensen, J. J., 1963. Factors affecting resistance of wheat to scab caused by Gibberella zeae. Phytopathology 53 7, 1: 831 - 838.Unger, P. W., 1994. Residue production and uses–an introduction to managing agricultural residues. In Managing Agricultural Residues; Unger, P. W., Ed., Lewis Publishers: Boca Raton, F. L., USA, pp. 1 – 6.Zhang, W., Boyle K., Brûlé - Babel, A. L., Fedak, G., Gao, P., Robleh Djama, Z., Polley, B., Cuthbert R. D., Randhawa, H. S., Jiang, F., Eudes, F., Fobert, P. R., 2020. Genetic Characterization of Multiple Components Contributing to Fusarium Head Blight Resistance of FL62R1, a Canadian Bread Wheat Developed Using Systemic Breeding. Front. Plant Sci. 11:580833.Zrcková, M., Svobodová - Leišová, L., Bucur, D., Capouchova, I., Konvalina, P., Pazderu, K., Janovská D., 2019. Occurence of Fusarium spp. In hulls and grains of different wheat species, Romanian Agricultural Research, No. 36, 173 - 185.Watkins, J. E., Boosalis, M. G., 1994. Plant disease incidence as influenced by conservation tillage systems. In Managing Agricultural Residues; Unger, P. W., Ed. Lewis Publishers: Boca Raton, F. L., USA, 261 – 283.Wegulo, S. N., 2012. Factors influencing Deoxynivalenol accumulation in small grain cereals, Toxins, 4, 1157 - 1180.Wang, Y. Z. and Miller, J. D., 1988. Screening techniques and sources of resistance to fusarium head blight. In: A. R., Khlatt, (ed), Wheat production: constraints in tropical environments. CIMMYT, Mexico. 239 - 250.***. 2006. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs.***. 2013. Commission Recommendation 2013/165/EU of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products.***. 2024. REGULAMENTUL (UE) 2024/1022 AL COMISIEI din 8 aprilie 2024 de modificare a Regulamentului (UE) 2023/915 în ceea ce privește nivelurile maxime de deoxinivalenol în produse alimentare, Jurnalul Oficial al Uniunii Europene, 9.4.2024, ELI: http://data.europa.eu/eli/reg/2024/1022/oj.
Articol scris de: dr. ing. OTILIA COTUNA, șef lucrări Facultatea de Agricultură USV „Regele Mihai I” Timișoara, Departamentul de Biologie și Protecția Plantelor
Foto: Otilia Cotuna (realizate în anii trecuți)
Abonamente Revista Fermierului – ediția print, AICI!
- Start
- Prec
- 1
- 2
- 3
- Mai departe
- Sfârșit

Publicitate







Revista
